An Efficient Mathematical Model Predicting the Number of Butterfly Species on Greek Islands in the Eastern Aegean
Hristos T. Anastassiu1* and Nikos Ghavalas2
1Department of Computer, Informatics and Telecommunications Engineering International Hellenic University, Serres Campus, End of Magnisias Str., GR-62124 Serres, GREECE
230 Karaoli-Dimitriou Street, GR-12461 Athens (Haidari), GREECE
Received Date: September 03,2025; Accepted Date: September 10, 2025; Published Date: September 25, 2025;
*Corresponding author: Hristos T. Anastassiu, 1Department of Computer, Informatics and Telecommunications Engineering International Hellenic University, Serres Campus, End of Magnisias Str., GR-62124 Serres, GREECE, Phone: +30 23210 49376, Email: hristosa@ihu.gr
Citation: Anastassiu HT, Ghavalas N (2025) An Efficient Mathematical Model Predicting the Number of Butterfly Species on Greek Islands in the Eastern Aegean. Adv Agri Horti and Ento: AAHE-228.
DOI: 10.37722/AAHAE.2025201
Abstract
Several computational models have been published in the literature designed to perform an a priori estimation of the number of butterfly species on islands. Typically the number of species is expected to increase with the island area A, and simultaneously to decrease with the minimum distance D from the closest mainland. The least complicated mathematical functions meeting these requirements are positive powers of A and negative powers of D. Taking into account documented numbers of species on a few pivot islands, the associated power constants are calculated and a regression line is finally drawn. In several cases such models prove to be insufficiently accurate, either because they are not specific enough, rendering them unable to simulate excessively large geographical regions, or because the proposed mathematical function is computationally unstable for very small D, since the denominator of the ratio gradually vanishes. In this paper, a simple mathematical model based on elementary linear algebra concepts is proposed for the special case of Greek islands in the Eastern Aegean. Using already known data for four islands only, judiciously chosen, a 4*4 linear system of equations is formulated, the numerical solution of which yields the power constants required to determine the appropriate mathematical expression. Moreover, negative exponential functions instead of negative powers are alternatively utilized for the D dependence, to circumvent the troublesome singularity at zero. Prediction results demonstrate very good agreement with actual data, proving the accuracy and versatility of the method.
Keywords: butterflies, exponential functions, Greece, islands, linear algebra, monotonic functions, prediction model
Watch the Article in Motion
Youtube shorts:
All the links shall be updated by Monday
Introduction
Islands all over the world are of great interest to Biogeography researchers, due to the exceptional conditions often dominating their autonomous environment. Isolation from the mainland gives rise to unusual behavior of their terrestrial flora and fauna; the species variability may differ significantly from the neighboring continental land mass, an effect that may be attributed to several reasons. Species continuously adapt in order to exploit the pertaining niche (quality of the environment), thus modifying the exploitation pattern of the latter [1]. This procedure often leads to endemism, which is a typical consequence of severe isolation. On the other hand, the number of non-endemic species, when compared to the closest mainland tends to be lower, predominantly due to unsuccessful adaptability and resulting extinction. According to [2], the actual number of species on an island is the outcome of a sequence of dynamic interactions between the rates of immigration and extinction.
There are several studies in the literature aiming to construct mathematical models that could possibly provide a pattern followed by the number of species on groups of islands. If the resulting pattern is adequately reliable, a priori predictions can be carried out before actual counts are performed in situ. Based on common sense, it is reasonable to assume that the larger the area A of the island, the richer its fauna/flora will be. Similarly, the greater the distance D from the closest mainland, the poorer the variability should be, due to increasing isolation [3]. Such considerations result into the following, initially abstract mathematical model: let N be the number of species of a particular animal group on a specific island. Since N must be an increasing function of A and a decreasing function of D, the simplest possible mathematical expression simulating such behavior is:
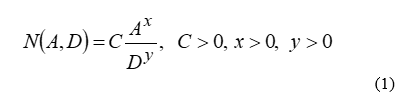
where C, x and y are positive constants to be somehow determined, as explained in the following sections. Although equation (1) is the basis of a large number of published papers on the topic of island biogeography, applications to butterflies are limited, whereas purely mathematical considerations thereof are seldom discussed. For one thing, (1) is often oversimplified, neglecting the effect of D (the classical Arrhenius model[4]). Furthermore, taking the logarithm of (1) is a common strategy, without clearly explaining the necessity of such a step. Sometimes the logarithm of A itself is assumed to be proportional to N (Gleason model [5]). In addition to this, dependence on extra variables, such as island maximum altitude or coordinates is taken into account heuristically, without mathematical rigor. Finally, eq. (1) and its variations are routinely utilized merely as input to commercial statistical computer tools that automatically produce “best fit” curves without any insight into the actual physical or mathematical mechanisms governing the evolutionary process.
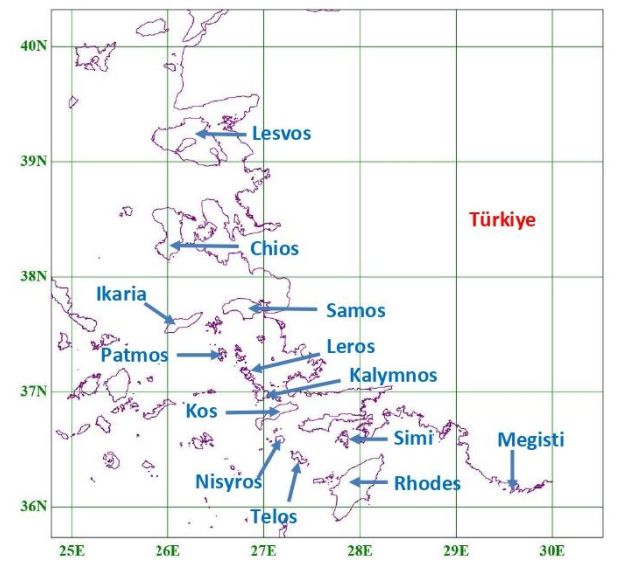
The aim of the present paper, which is an extended version of [6], is two-fold: its first, obvious, aspect with practical use is to propose an improved mathematical model, accurately predicting the number of butterfly species on Greek islands in the Eastern Aegean. Moreover, the second and most important one is to demonstrate how such a model can be derived in a fairly simple and straightforward manner from a minimal amount of data, associated with only four islands, without the need of statistical investigation into extensive data sets. Although the model can be applied to other island groups, Eastern Aegean islands were deliberately chosen as a challenging case study for various reasons: they share a common closest mainland, they demonstrate great variability in terms of A and D and they form some sort of bridge between Europe and Asia, combining elements of the fauna encountered on both continents.
Earlier Models And Their Applicability
Although the literature on island biogeography is very extensive, applications to butterflies are rather scarce. As discussed earlier, [2] proposed a model for the butterflies of the Mediterranean islands. The mathematical analysis presented in [2] is somewhat obscure; originally, (1) is used as the fundamental equation without the involvement of D (in other words, y is implicitly set equal to 0). After constructing a corresponding regression line using 12 islands of the Mediterranean, all the sudden D is heuristically added to the procedure, and a multiple regression analysis is performed, resulting in the following “best fit” expression:
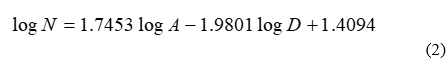
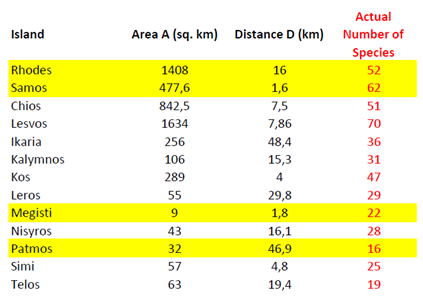
where A is expressed in square miles, D in miles and log denotes the base-10 logarithm. The author discusses potential limitations of this model, especially when islands are clumped together or are connected to the mainland by a series of stepping stones. To assess the accuracy of (2) in analogous situations, a few calculations were carried out by our research team for several Greek islands in the Eastern Aegean (Fig. 1). Results are presented in Table 1, obviously showing an exorbitant deviation from actual values. Like pointed out above, [2] warns for potential cases where (2) may be inaccurate, however in this particular island group the numbers predicted are orders of magnitude different than the actual ones. Therefore, the following question is raised: why does eq. (2) fail so emphatically for the Eastern Aegean, since it is apparently satisfactory for the 12 Mediterranean islands chosen in [2]?
There are several sources of this severe discrepancy. The most important one is that the 12 islands considered in [2] cover an excessively large area, with group members of completely dissimilar properties, such as Sardinia and Karpathos. From a mathematical point of view, this is bound to lead to failure for the following reason: the C constant in (1) obviously depends on the associated butterfly fauna of the closest mainland shore. However, in [2], the closest mainland is not the same for all islands considered (it may be Spain, Italy, Greece or Türkiye). Therefore, it is not possible to define a unique, suitable C for the entire group of islands described in [2], and hence (1) and (2) cannot possibly be valid.
Moreover, the occurrence of D in the denominator of (1), and in the logarithm argument in (2), causes additional problems. From a mathematical point of view, both (1) and (2) are singular at zero, meaning that D cannot possibly be set equal to 0, because N would then tend to infinity. Of course, in realistic situations, D is never equal to 0, because in that case the land mass considered would not be an island. However, if D is very small, it could still cause possible numerical instabilities, especially if the y constant (or, equivalently, the constant multiplying log D in (2)) has not been carefully determined. Such numerical instabilities are obvious in Table 1, where all islands investigated lie fairly close to the opposite Turkish mainland (see Fig. 1).
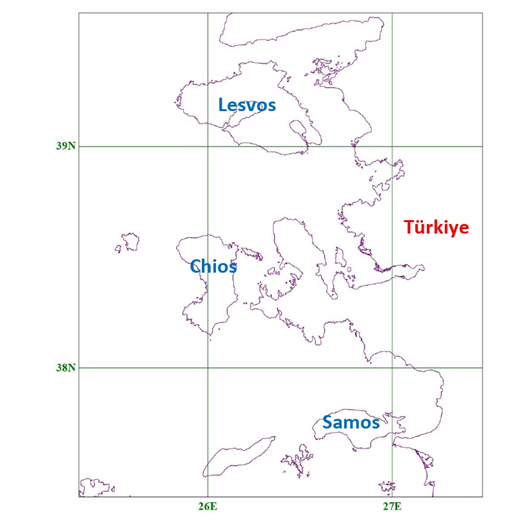

Further research works dedicated to the Aegean islands are found in [7,8]. Rather than developing a mathematical model and presenting its derivation, the analysis performed is purely statistical. A considerable number of islands are tabulated (31 in [7] and 64 in [8]), while multiple linear stepwise regression is performed, based on 9 geographical variables, including, in addition to A and D, maximum altitude, both distances from Greek and Turkish mainland, coordinates, isolation measure e.t.c. Numerical results are apparently extracted via some computational tool without any details on the incorporation of the extra geographical variables into (1). Analogous concepts have been extended to the totality of European islands in [9], again with emphasis in Statistics, rather than modeling.
A Novel Mathematical Model
In order to develop a new, simple, yet efficient mathematical model for the distribution of butterfly species on Greek islands, (1) is assumed to be fundamental in the analysis, since it provides the simplest mathematical expression possible that meets the criteria discussed in the Introduction. The actual relationship between N and the independent variables, especially A, has been a matter of dispute for more than a century [10]. Similarly, the dependence on D should be simulated in a very careful way, for reasons explained in Section 2. To mitigate potential instabilities, the following modification of (1) is proposed herein:
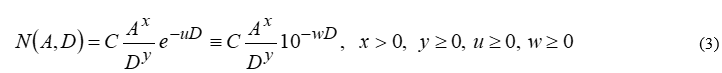
i.e. an exponential term associated with D is assumed to multiply the ratio in (1), making the composite model more flexible and versatile. The two forms in (3) are equivalent, since w=u log e. Note that the y, u and w constants are theoretically allowed to vanish; if y=0 , the function becomes exclusively exponential, whereas whenu=w=o, (1) is recovered. Therefore, (3) involves four unknown constants, i.e. C, x, y and w which must be determined. To this end, a linear system of equations may be constructed, where these four quantities are unknowns; to attain linearity, the logarithm of (3) is taken. This is the reason for the widespread use of the logarithm (and not the assertion that large numbers come into play, as usually believed). The outcome yields:
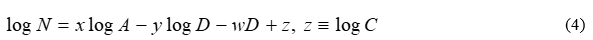
The units for A and D do not matter, as long as their use is consistent. If (4) is applied to four pivot islands with known number of species, i.e. N1 N2, N3, N4 respectively, the unknown constants are equal to the solution of the 4*4 linear system formulated, i.e.:
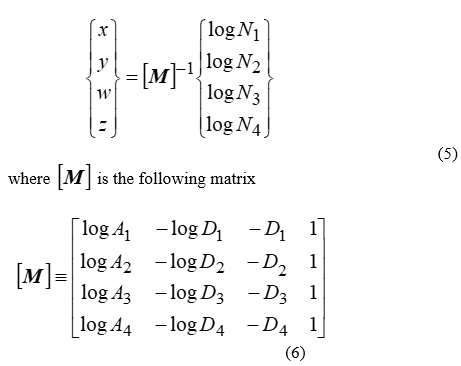
and [M]-1 is the inverse of [M]. Four is the minimum number of islands that should be used in the aforementioned procedure however taking into account even more is possible. If Q>4 islands are tabulated and utilized as a pivot, then an over-determined linear system is constructed, which does not necessarily have an exact solution. The “best-fit” solution, i.e. the solution accompanied by the lowest possible error, is given in terms of the Moore-Penrose pseudo-inverse of [M], the latter retaining its form in (6), but now having Q rows:
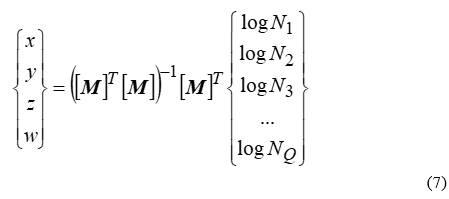
where [M]T is the transpose of [M]. This is exactly the core of multiple regression analysis which is a well-known tool in Statistics.
Numerical Results and Discussion
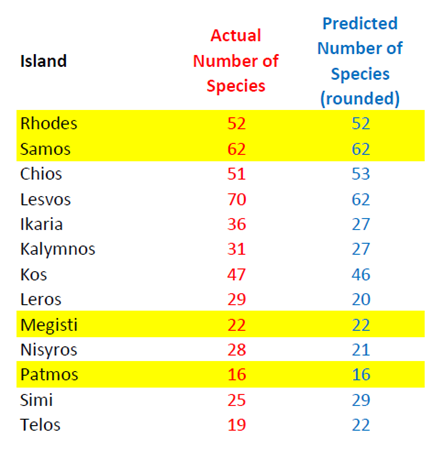
To test the accuracy of the model presented in Section 3, the simplest possible scenario was introduced for a wide selection of Eastern Aegean Islands (Fig. 2). The minimum amount of pivot islands, i.e. 4, was used to assess the reliability of the model for the lowest amount of resources available. To cover the broadest spectrum of cases, the pivot choice made included both large and small islands, close and far away from the coast. A suitable group is: Samos (large and close), Rhodes (large and distant), Megisti (small and close) and Patmos (small and distant). By transferring the corresponding values from Table 2 to (5) and (6), and solving the system, the following complete expression is derived for (4):
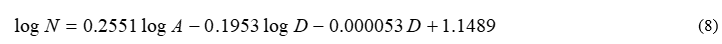
where A is expressed in square km and D in km. Using (8) for the remaining islands of the group yields the results shown in Table 3. Evidently, the model derived in (8) demonstrates remarkable accuracy for almost all cases, although only four islands were used as a pivot. Using a higher Q in (7) is allowable, and (7) would minimize the total error, which is a well-known theorem in Linear Algebra. However, this is not the point of this paper; the main purpose is to prove that a reliable model can be constructed, without the requirement of large amounts of accessible data for many islands. Indeed, just four of them are proven to be sufficient.
Investigation of the coefficients in (8) draws additional conclusions; as discussed in [10] it is not generally clear if models of this type should follow a power or exponential behavior, causing a lot of disputes in the research community. The mathematical procedure described above permits the answer to this problem to develop automatically: indeed y expresses the power dependence and w the logarithmic one. Solution of the linear system in our case yields y=0.1953 and w= 0.000053 (see (8)), meaning that the exponential behavior is almost negligible herein (but not necessarily so in other analogous situations, where D may approach 0).
Compared to analogous models in recent literature, (4) is formally similar to the basic expression in [11], however there are still important differences. In [11] the number of niches is assumed to be greater than 1, and the species richness is assumed to vary with time, whereas this work includes the novel exponential term that was previously discussed in detail. In [12] there is an extensive literature collection, however, as admitted therein, “historically, there have been fewer applications of Species Distribution Models in island environments and for island species”. Furthermore, in recent literature emphasis is put mainly on Statistics of large data sets [13], and not on Mathematics, making any direct comparison almost impossible.
Refinement of the model may include addition of extra variables, such as the ones used in [7,8]. It is particularly interesting to investigate the effect of maximum altitude, since several islands in the region possess relatively high mountains rising up to 1433 m (e.g. Mt. Kerkis on Samos). Future work is planned to incorporate this information to a possible model extension.
Summary And Conclusions
In this paper, a mathematical model predicting the number of butterfly species on Greek Islands in the Eastern Aegean was derived from completely basic concepts. Assuming a particular type of dependence on variables such as island area and distance from the closest mainland, four key constants were determined though the solution of a linear system of equations. Although a larger number of islands with known fauna could be used, only four of them, judiciously chosen, were proven to be sufficient for the construction of a reliable model, meaning that it is not imperative to statistically process a large amount of available data, which is the norm in most publications found in literature. This model may be utilized for several practical applications such as biodiversity monitoring, even for other island groups or even for other taxa. If severe discrepancies are observed between predicted and actual number of species, especially if the latter are much lower than expected, conservation planning should probably take effect, to prevent possible further impoverishment of the local biotopes. Assessment of the ecological impacts of climate and land-use changes, and biological invasions may be another field of application [12].
Author Contributions
Hristos T. Anastassiu: mathematical analysis, calculations, manuscript preparation
Nikos Ghavalas: original concept, literature search
References
- Root RB (1967) The niche utilization pattern of the blue gnatcatcher. Ecol. Monogr. 37: 317-350
- Hocking DC (1980) The biogeography of the butterflies of the Mediterranean islands. Nota Lepid. 3 (3-4): 119-125
- Mayr E (1940) The origin and history of the bird fauna in Polynesia. Proc. 6th Pacific Science Congress 4: 197-216
- Arrhenius O (1921) Species and area. Journal of Ecology. 9: 95-99.
- Gleason HA (1922) On the relation between species and area. Ecology, 3: 158–162
- Anastassiu HT and Ghavalas N (2025) An Improved Mathematical Model Predicting the Number of Butterfly Species on Greek Islands. 16th International Congress on the Zoogeography and Ecology of Greece and Adjacent Regions (ICZEGAR), Athens, Greece
- Dennis RLH, Shreeve T, Olivier A and Coutsis JG (2000) Contemporary geography dominates butterfly diversity gradients within the Aegean archipelago (Lepidoptera: Papilionoidea, Hesperioidea). Journal of Biogeography. 27: 1365-1383
- Dennis RLH, Olivier A, Coutsis JG and Shreeve T (2001) Butterflies on the islands in the Aegean archipelago: predicting numbers of species and incidence of species using geographical variables. Entomologist’s Gazette 52: 3-39
- Dennis RLH et al. (2008) Butterflies of European islands: the implications of the geography and ecology of rarity and endemicity for conservation. J. Insect Conserv. 12: 205-236
- [10] Tjørve E (2012) Arrhenius and Gleason revisited: New hybrid models resolve an old controversy. Journal of Biogeography. 39(4): 1-11
- Beaugrand G, Kléparski L, Luczak C, Goberville E and Kirby RR (2024) A niche- based theory of island biogeography. Ecology and Evolution 14:e11540. DOI: 10.1002/ece3.11540
- Rios EB, Sadler J, Graham L, Matthews TJ (2024) Species distribution models and island biogeography: Challenges and prospects. Global Ecology and Conservation. 51:e02943
- Shuchan Zhou, Kan Wang, Beata Messyasz, Yaoyang Xu, Meixiang Gao, Yuying Li, and Naicheng Wu (2022) Functional and taxonomic beta diversity of butterfly assemblages in an archipelago: relative importance of island characteristics, climate, and spatial factors. Ecological Indicators, 142: 109191
Appendix: