An Excursus on the Role of Wildlife Carcasses
Claudia Jordan-Fragstein1*, Thomas Hofmann2, Christian von Hoermann2 and Michael Müller1
1Chair of Forest Protection, Technische Universität Dresden, Tharandt, Saxony, 01737, Germany
2Civil Service Trainee in Forestry Administration, Saxony Forestry Department, Germany
3Chair of Wildlife Ecology and Management, Albert-Ludwigs-Universität Freiburg, Germany
Received Date: May 26, 2025; Accepted Date: June 18, 2025; Published Date: June 26, 2025;
*Corresponding author: Claudia Jordan-Fragstein, Chair of Forest Protection, Technische Universität Dresden, Tharandt, Saxony, 01737, Germany; Tel: +49 15155624424; Fax: +49 351 463-31397; Email: claudia.jordan-fragstein@tu-dresden.de
Citation: JaFragstein JC, Hofmann T, Hoermann CV, Müller MS (2025); An excursus on the role of wildlife carcasses Enviro Sci Poll Res and Mang: ESPRM-171
DOI: 10.37722/ESPRAM.2025202
Abstract
Carrion represents a highly dynamic and nutrient-rich resource in terrestrial ecosystems, yet its ecological role remains underrepresented compared to deadwood. This study investigates the necrophilous beetle communities (Coleoptera) associated with roe deer (Capreolus capreolus L.) carcasses in a Central European forest ecosystem. Four carcasses were placed in two forest habitats (coniferous and deciduous) during summer and winter seasons in the Tharandt Forest (Saxony, Germany). Arthropod succession was systematically recorded over several weeks using pitfall traps, direct sampling, and climate monitoring. The results revealed distinct seasonal and habitat-related differences in species composition and abundance. Silphinae, Staphylinidae, and Histeridae dominated the winter assemblages, while Dermestidae and Cleridae became prominent in later summer decomposition stages. Seasonal variation in temperature and microhabitat influenced the rate and pattern of carcass decomposition and arthropod colonization. The study also confirmed that larger carrion remains, such as roe deer, support a broad spectrum of trophic guilds and provide habitat for both obligate and facultative necrophilous beetles. The findings emphasize the importance of medium-sized ungulate carcasses in maintaining coleopteran diversity and underscore their potential role in supporting ecosystem functions such as nutrient cycling and food web stability. Given the increasing anthropogenic reduction of carrion in managed landscapes, we recommend considering natural carcass availability in biodiversity conservation strategies. This study contributes to the growing field of carrion ecology and provides a methodological framework for long-term monitoring of necrophilous insect diversity in forested environments.r superior source for bioethanol, compared with sugar beet.
Keywords: carrion ecology, Coleoptera, decomposition, forest biodiversity, insect succession, roe deer, necrophilous beetles, ecosystem services
Watch my article in motion:
Information Board
ISSN: 2641-6816
Frequency: Continuous
Format: Video, PDF and HTML
Versions: Online (Open Access)
Language: English
Impact Factor: 4.21
Quick Links
Issues
Video Gallery
Guidelines
Publication Fee
Editorial Board
Review Board
Publish your article not just in PDF and HTML, but also in documentary format. Simply submit your manuscript as usual — we’ll take care of the rest.
Introduction
Carrion is a central yet often overlooked resource in terrestrial ecosystems. As a highly nutrient-rich substrate, it provides energy and biomass for a diverse array of specialized and generalist organisms, thereby playing a vital role in nutrient cycling, habitat provision, and the promotion of functional biodiversity. Despite this ecological relevance, the role of vertebrate carcasses has received little attention in forestry and conservation strategies—especially when compared to the well-established ecological functions of deadwood (Barton et al. 2013b). Numerous organism groups depend on carrion, particularly insects with necrophilous life strategies.
Coleoptera (beetles) represent a highly diverse and functionally important group in this context. They utilize carcasses in various trophic roles—as necrophages, predators, parasitoids, or detritivores. Their succession follows typical decomposition stages, which are in turn influenced by carcass condition, size, microclimate, and access by other scavengers (Payne 1965; Goff 2009; Matuszewski et al. 2010a). While such succession patterns are well documented in forensic entomology, ecological field studies on larger vertebrate carcasses in Central European forests remain scarce.
The roe deer (Capreolus capreolus L.) is the most abundant wild ruminant species in Germany, with over one million individuals legally harvested each year (DJV 2022). Its broad distribution and high population density make it a suitable model organism for carrion ecology. Roe deer carcasses are large enough to attract a wide range of decomposer insects and occur in both deciduous and coniferous forest types. In practice, however, most carcasses are removed from the forest due to hunting regulations and sanitary concerns. This includes game animals as well as roadkill and other mortality events, leading to a significant reduction in carrion availability for necrophilous organisms.
Carrion availability is also subject to seasonal variation. In summer, decomposition proceeds rapidly under high insect activity, whereas in winter, decomposition is delayed and insect succession patterns are markedly different. Habitat type further modulates these dynamics: factors such as microclimate, soil structure, and canopy cover can significantly influence species composition (Matuszewski et al. 2010a; Parmenter & MacMahon 2009). Anthropogenic influences, such as forest management, hunting practices, and habitat fragmentation, may also alter carrion dynamics and its ecological functions (Anderson 2020).
Carrion ecology is an emerging subdiscipline that integrates aspects of decomposition, biodiversity, and resource dynamics. However, empirical data on the arthropod communities associated with large mammal carcasses under variable environmental conditions remain limited. In particular, comparative studies across seasons and forest types are lacking, as are data on the ecological significance of carcasses for beetle communities in temperate forest systems.
This study addresses these research gaps by investigating the necrophilous Coleoptera associated with four roe deer carcasses in the Tharandt Forest (Saxony, Germany). Two forest habitats a beech-dominated deciduous stand and a spruce-dominated coniferous stand were selected, with carcasses deployed in both summer and winter. The study aims to document arthropod succession, diversity, and community structure over time, assess seasonal and habitat-based differences, and explore implications for forest biodiversity and ecosystem function. In doing so, it contributes to the recognition of carcasses as functional ecological resources in forest ecosystems and provides a methodological basis for further research in carrion ecology.
The conceptual diagram in Figure A illustrates the complex trophic interactions and nutrient flows triggered by the introduction of an animal carcass into a terrestrial ecosystem. Carcasses act as highly concentrated sources of energy and nutrients and play a central role in structuring local material flows and food webs (Bump et al., 2009; Benbow et al., 2019).Immediately after the death of an organism, initial decomposition occurs through scavengers (e.g., vertebrates such as birds and mammals), which remove coarse tissue and mobilize large amounts of biomass. In the second phase, detritivores (e.g., insect larvae, earthworms) and microbial decomposers (e.g., bacteria, fungi) take over the further decomposition of organic matter. These organisms catalyze the mineralization of the carcass, releasing bioavailable nutrients, especially nitrogen compounds and phosphorus, into the surrounding soil (Janzen, 1977; Barton et al., 2013). The soil nutrients enriched in this way promote the growth of primary producers, allowing locally productive hotspots to develop within the ecosystem. These vegetation-rich zones in turn provide an improved food source for herbivorous organisms and thus directly influence higher trophic levels. Ultimately, secondary and tertiary consumers, such as predators, also benefit indirectly from the trophic cascade induced by the carcass (Wilson & Wolkovich, 2011; Carter et al., 2007).These ecological feedback effects show that carcasses should not be viewed merely as the end points of individual life cycles, but rather as integral components of dynamic material cycles that substantially influence biodiversity, nutrient availability, and energy flows within ecosystems.
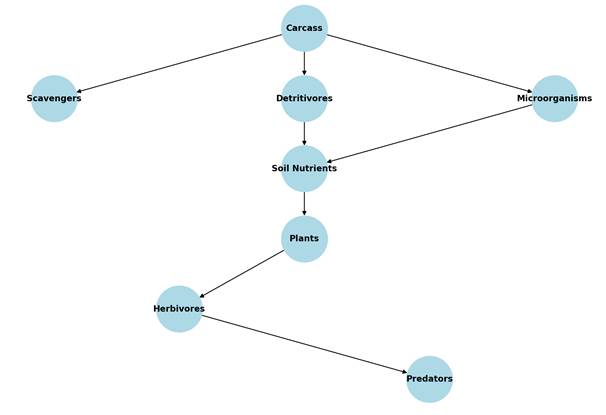
Figure A: Trophic interactions and Nutriens Pathways from Carcass
Method
General Research design
Experimental Environment
The study site is located in the Tharandt Forest, within the wildlife enclosure of TU Dresden, in the state forest district Grillenburg, compartments 340 and 341 of the Bärenfels Forest District. The wildlife enclosure lies south of the E-wing of the university buildings and is bordered to the east by the Grunder Weg. The climate of the Tharandt Forest corresponds to that of the lower montane zone of the Ore Mountains (GOLDBERG et al. 2002). In this region, two macroclimate types converge: a submontane climate represented by the “Klingenberg” climate type and a climate influenced by the Elbe valley, represented by the “Glashütte” climate type (SAXON STATE OFFICE FOR ENVIRONMENT, AGRICULTURE AND GEOLOGY 2010). According to the nearby climate station in Grillenburg, the long-term average annual temperature is 7.8 °C, and the average annual precipitation is 901 mm (CHAIR OF METEOROLOGY, 23 January 2021). The prevailing soil types are podzolic brown earth and a pseudo-gley affected by stagnant water (LFULG & SMEKUL 2022).
The wildlife enclosure covers approximately 9 hectares and is home to red deer (Cervus elaphus L.). In the following, these animals are referred to as “red deer.” They are kept under near-natural conditions to allow for the study of natural behaviors (INSTITUTE OF FOREST PROTECTION). At the time of the experiment, trials on acoustic deterrence of red deer were being conducted. The density of animals—four individuals within the enclosure—is considerably higher than in natural forest environments. Due to the increased amount of dung, the enclosure represents a special experimental environment for a study of this kind.
Selection of Forest Habitats
To compare carrion colonization between two forest types, sites within the wildlife enclosure were selected that typically represent a coniferous stand and a deciduous stand, respectively.
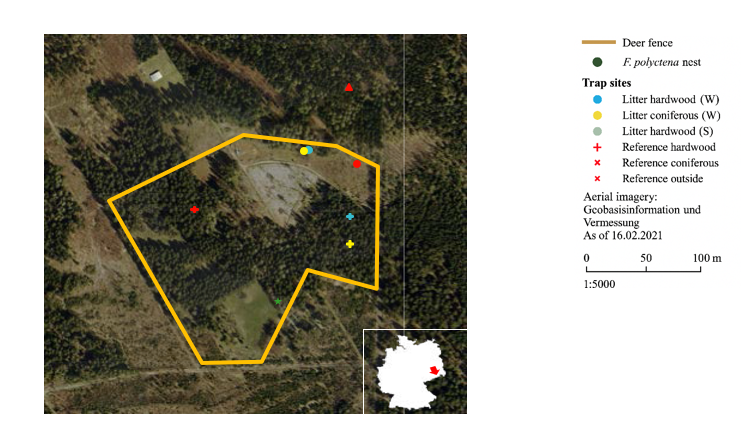
Fig. 1: Experimental sites; abbreviations W and S indicate the experimental periods Winter and Summer, respectively.
A closed pure stand of Norway spruce (Picea abies H. KARST.) with occasional individual admixture of silver birch (Betula pendula ROTH) was selected as the coniferous forest type. This stand is estimated to be approximately 80 years old. Norway spruce is the most common main tree species in Saxony but has been significantly affected by recent drought years and bark beetle outbreaks (SMEKUL 2022). Larger deciduous stands are not present within the wildlife enclosure, and fenced-off areas with black alder (Alnus glutinosa L.) in the thicket stage were considered less suitable. Reasons include both the surrounding fence, which excludes red deer, and the characteristics of the stand itself. The area is still in an early developmental stage, and due to its tree species composition, it is only representative of potential natural vegetation (pnV) in a few locations of the Tharandt Forest.
As a deciduous forest stand, a small area dominated by European beech (Fagus sylvatica L.) was selected, featuring a centrally located, large-diameter veteran beech with a broad crown. Along the adjacent path to the south, five younger beech trees in the pole wood stage are present. As shown in Figure 1, the sampling sites within the deciduous stand are located in close proximity to each other.
According to vegetation mapping, the potential natural vegetation (pnV) at the experimental site consists of acidophilous beech(-mixed) forests in various forms (LFULG & SMEKUL 2022). The selected stand composition therefore closely resembles the local pnV.
Due to the influence of red deer and the resulting heavy shading, both selected forest stands exhibited almost no ground vegetation. Accordingly, no vegetation survey was conducted. Since 2018, repeated salvage logging operations due to calamities have taken place within the enclosure. The resulting open area borders the selected beech stand and influences the microclimatic conditions of the deciduous site. In contrast, the coniferous experimental sites are located at a greater distance from the open area.
Three types of reference sites were selected. First, a comparable section of the coniferous stand in the northwest of the enclosure was designated as a reference. For the deciduous stand, the area of black alder in the thicket stage located to the east was used as a reference. A third reference site was established outside the enclosure to exclude the influence of the high red deer density. This site, situated adjacent to an unused former enclosure north of the facility, consists of a spruce-dominated mixed stand with advance growth of silver fir (Abies alba MILL.), discontinuous canopy closure, diverse natural regeneration, and herbaceous ground vegetation. The fence line shown on the map represents the boundary of the red deer enclosure. Other enclosures are currently not in use. For improved readability of the study, the carcass placed in the coniferous forest is referred to as Carcass Conifer, and the carcass in the deciduous forest is referred to as Carcass Deciduous. These designations apply consistently across both experimental periods.
Carcasses Used
In this study, complete animal carcasses are deployed to provide a natural diversity of resources for necrophilous arthropods. In accordance with the principles of carrion ecology, wild animals are used that are free from pharmaceutical contamination, fed on natural diets, and possess fur typical of wild mammals (GU et al.). The roe deer, a common medium-sized vertebrate, is selected as the model species. To reflect the potential spectrum of necrophilous species associated with large carcasses, the roe deer should have a target minimum weight of approximately 20 kg (HÖRMANN 2021).

Fig. 2: Female roe deer (left) and male roe deer (right) in a freshly dead state, placed in the mesh cage at the start of the experiment on 15 January 2021. The arrow indicates the location of the gunshot wound.
The carcasses used in the winter trial consisted of two shot animals. A multiparous female roe deer weighing 21.8 kg and a multiparous male roe deer weighing 26.2 kg were used. Thus, the larger individual was approximately 20% heavier than the smaller one. Both animals died instantly. The female was shot with what is referred to in hunting terminology as a “high lung shot” (“Hochblattschuss”), with the entry and exit wounds located just behind the shoulder blade. The bullet struck the lungs a few centimeters below the spine, resulting in immediate death without flight. The male roe deer was killed with a “neck shot” (“Trägerschuss”) at close range, which hit one of the lower cervical vertebrae. This caused injury to the spinal cord and likewise led to immediate death. Consequently, both animals died without suffering and without attempting to flee. For the study, a relevant difference between the two carcasses is that the thoracic cavity of the female was already opened by the shot canal. The exit wounds, facing downward in the experimental setup, measured approximately 3.5–4.0 cm in the female and around 2 cm in the male.

Fig. 3: Yearling female roe deer (Schmalreh) killed in a traffic accident (left) in winter coat, and shot Schmalreh (right) in summer coat, both placed at the experimental site in a freshly dead state on 5 August 2021 after being deep-frozen.
For the summer trial, roe deer killed in road traffic accidents—referred to in hunting terminology as “fall game” (“Fallwild”)—were intended to be used exclusively. After one such specimen was successfully obtained, all further efforts by the author and the institute to acquire a comparable roe deer body proved unsuccessful. In order to conduct the experiment within the planned timeframe, a second roe deer was therefore shot. n both cases, the animals were yearling female roe deer, referred to in hunting terminology as Schmalreh (see Figure 3). The first individual died in a traffic accident on 18 February 2021 and weighed 17.9 kg. The skull was fractured in multiple places, and several broken ribs were present. Additionally, the skin on the left foreleg was torn. It is assumed that the animal died immediately upon impact. Prior to deep-freezing, bloating of the rumen area had already begun. The second roe deer was shot on 3 August 2021 and weighed 21.6 kg. Again, the weight difference between the two animals is approximately 20%, relative to the lighter individual. The bullet path in the second deer was located in the posterior lung area, resulting in death after a short escape distance. Upon closer inspection of the carcass, an old fracture of the right foreleg was detected, which is considered irrelevant to the experimental course. Since the first roe deer had already been stored at –28 °C for several months, and freezing inevitably delays decomposition (PAYNE 1965; ROBERTS & DABBS 2015; STIEGLER et al. 2020), the second, freshly shot animal was also deep-frozen at –28 °C for 48 hours. This ensured that both carcasses were equally frozen prior to the experiment. Due to the different seasons at the time of death, the first roe deer still bore its winter coat, while the second was in summer coat in early August. At the start of the experiment, both animals were weighed in their frozen state and directly placed into the mesh cages at the designated new locations.
Experimental Setup and Timeline
Two roe deer carcasses were deployed during each trial period—on 16 January 2021 in winter and on 5 August 2021 in late summer. To protect the carcasses from large vertebrate scavengers, particularly foxes and wild boars, they were enclosed in steel mesh cages. This measure was primarily intended to prevent the loss of substantial carcass material during the winter and to avoid significantly affecting arthropod colonization. The cages were anchored with ground stakes. On the short sides of each cage, additional steel T-profiles were driven into the ground and fixed to the cages. These T-profiles, in combination with a wooden slat, were used to repeatedly position a smartphone camera (15.9 megapixels, aperture f/1.9) during sampling events.
To minimize disturbance to the soil structure, no digging protection or similar barrier was installed. Small birds and mustelids were consistently able to enter the cages throughout the experiment.
To capture arthropods around each carcass, eight BARBER traps were installed per site (BARBER 1931) (see Figure 4). These consisted of honey jars (capacity 405 ml) inserted flush with the soil surface and fitted with a screwed-on cover plate serving as a walking surface. Each jar was filled with a solution of saturated benzoic acid and household dish detergent to reduce surface tension. Typically, 100 ml of trapping liquid was used to facilitate later decanting and to prevent larger Coleoptera from reaching the rim even when rainfall increased the fluid volume. In periods of high arthropod activity, the volume of trapping liquid was increased to ensure sufficient capacity for specimen collection.
The jars were held in place solely by the surrounding cover plate at the soil surface. No internal plastic sleeve was used to prevent soil from collapsing into the jar. This design choice was made so that arthropods trapped beneath the round cover plate and under the jar itself would have a better chance of escaping.
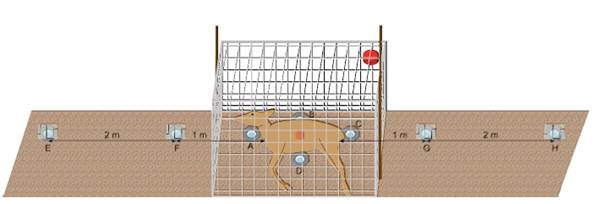
Fig. 4: Schematic representation of the experimental setup
For the experimental setup, four BARBER traps were placed inside the cage in an inner radius around the carcass, and two traps each were positioned outside the cage at the front and rear ends of the body, at distances of 1 m and 3 m. Within the inner radius, the traps were installed in direct contact with the carcass. Their positions were at the head, mid-back and abdominal line, and the anus (cardial, ventral, caudal, lateral) and labeled A to D in a clockwise direction. To investigate potential effects on the surrounding arthropod fauna, traps were also deployed outside the mesh cage along the longitudinal body axis at 1 m and 3 m distances. These exterior traps were covered with acrylic roofs to prevent rain from diluting the trapping fluid or causing overflow. For practical reasons, no rain covers were used on the traps directly adjacent to the carcass. Any traps compromised by soil, rumen contents, or hair were initially recorded as “Category II.” However, this categorization was later abandoned, as contamination became increasingly unavoidable throughout the experiment. Excluding these traps from data analysis would have rendered the results incomplete in terms of catch numbers and trap positioning.
A data logger for ambient temperature recording was mounted in an upper corner of the cage at approximately 0.9 m height, protected by a small enclosure. This logger is indicated in red in the schematic. A second data logger was placed underneath the carcass torso to record the temperature between the soil surface and the carcass; it is shown in orange in the diagram. In addition to the setups at the carcass sites, reference trap arrays were deployed in the surrounding area. These consisted of six traps arranged along a single axis. Positions D and B were omitted in this configuration.
Sampling Periods
In the winter experiment, pitfall traps were installed later than planned due to persistent snow cover. At the deciduous forest carcass site, traps could be deployed on 18 February 2021 (day 40), as the snow had melted earlier in this area. At the coniferous forest site and on the reference plots, the traps were installed on 26 February 2021 (day 43). The trapping period for the deciduous forest carcass lasted from 23 February to 7 June 2021, and for the coniferous forest carcass from 26 February to 7 June 2021.
In the summer experiment, pitfall traps were installed at both experimental sites on 5 August 2021, with the first emptying occurring on 7 August 2021. The regular dismantling of the traps was scheduled for early September. At the coniferous forest site, traps were removed on 2 September 2021. However, at the deciduous forest carcass site, the trapping period was extended due to renewed high arthropod activity and the presence of residual muscle tissue, particularly in the lower hind limb region. As a result, the traps at this site remained active until 10 September 2021.
Manual Collection of Arthropods
In addition to emptying the deployed pitfall traps, arthropods were manually collected from the carcasses on each sampling day. This involved lifting the carcass to inspect both its surface and underside. The purpose of manual collection was to detect potentially rare species that are easily recognizable by eye early in the decomposition process, and to document characteristic species present at each specific time point. Manual collection also offers the possibility of detecting rare species that might not be captured by traps. As moving the carcass constitutes a disturbance, it was handled as gently as possible and subsequently repositioned in its original place. To ensure reproducibility and to minimize interference with arthropod succession, a fixed time window of three minutes per carcass and sampling day was applied. These samples are therefore referred to throughout this study as “3-minute samples.” Both Diptera larvae and Coleoptera were collected, although only a limited number of individuals were taken, in accordance with their observable abundance and morphological characteristics.
Measurement of Abiotic Factors
Temperature and Humidity
To measure temperature and humidity, data loggers of the type Tinytag Plus 2 (alternative designation: TGB-4500, Gemini Data Loggers UK Ltd.) were used. The positions of the data loggers (indicated in red and orange) are shown in Figure 5. Measurements were recorded hourly. After the experiment was concluded, the data were read out.
Placing these relatively large data loggers inside the carcass would have required invasive procedures and was therefore avoided. Additional climatic data were obtained from the nearby weather station in Grillenburg (Chair of Meteorology, 23 January 2021).
Indirect Radiation Measurement
To determine the radiation conditions around the carcass and the pitfall traps, hemispherical photography was used. For this purpose, a digital camera (model: Nikon Coolpix E995, 3.34 MP) equipped with a Nikon FC-E8 fisheye converter was mounted on a tripod at a height of 1 meter, leveled with a spirit level, and aligned to the north using a compass. Exposure time and aperture were manually set using a light meter (see Figure 24).
The method of hemispherical canopy photography and automated image segmentation was developed at TU Dresden for forest science applications, especially in tree regeneration ecology. It has been validated through practical trials and provides meaningful results even in low-light conditions with radiation levels below 10% of open-field radiation (SCHWALBE et al. 2009). In this method, shaded and unshaded image compartments are classified and segmented using in-house software from the Chair of Remote Sensing based on grayscale values, and then output as black-and-white negatives for manual verification. The user can manually refine the segmentation using this output (see Figure 25).
Based on the segmented images, the sun’s path is simulated over the image compartments, allowing calculation of ground-level radiation. In this study, the simulation of the sun’s path corresponds to the sun’s trajectory in June (days 150–170 of the year) in the Tharandt Forest, representing a maximum potential global radiation of 427.1 W/m² under clear-sky conditions. The global radiation at each trap location was calculated relative to this theoretical maximum during the simulated June sun path. Although actual radiation levels are significantly lower at the beginning of the year, the relative open-sky radiation values between sampling points remain comparable.
For traps A–D, which were arranged around the carcass, a central hemispherical image was taken above the carcass. To obtain an average radiation value for the reference plots, three hemispherical photographs were taken. Only the reference site in the deciduous forest habitat was photographed a second time at the end of the growing season. In this case, the camera was horizontally aligned 40 cm above the ground to account for the influence of bracken fern (Pteridium aquilinum).
Scavenging by Vertebrate Scavengers
In natural decomposition processes, carnivorous mammals play an important role in carrion utilization (source). Although the experimental setup excluded such animals through the use of protective cages, camera traps were used to document vertebrates that were attracted to the carcasses. Due to the fenced enclosure, large vertebrates such as wild boar (Sus scrofa L.) were largely excluded. Therefore, small to medium-sized carnivores were more likely to be recorded at the carcass sites.
Camera traps of the model Cuddeback C2 (aperture f/2.7) were used, with two cameras aimed at each experimental site from a distance of 3–4 meters. Only vertebrates in the immediate vicinity of the carcass setups were recorded. Red deer (Cervus elaphus) present in the enclosure were not included in the analysis. Observations regarding the potential repellent effects of roe deer carcasses on red deer movement were deemed unrepresentative, as the confined enclosure resulted in unnaturally high population densities and limited movement ranges. In addition, red deer activity near the deciduous forest setup was influenced by the proximity of the enclosure fence.
Animal visits recorded at the site ceased abruptly following repair of the inner fence. No clear statement can be made regarding the barrier effect of the outer fence for wild boars. In the past, wild boars repeatedly breached the fence, although it was regularly repaired to prevent this.
Identification of Coleoptera
Pitfall trap samples were cleaned by manually removing coarse soil particles, hair, and—in some cases—rumen content. If fine soil particles were present, the samples were rinsed using a fine mesh sieve. Frequently encountered Diptera larvae were separated for potential later quantification or identification. All samples were preserved in a 70% ethanol solution for long-term storage. Large quantities of samples, including separated Diptera larvae and entire bulk collections that could not be processed immediately, were temporarily frozen. In some cases, water was added prior to freezing to prevent desiccation and damage to exposed specimens. Subsequent identification of beetles was carried out under a stereomicroscope (Zeiss Stemi 508), with a focus on necrophilous species. All Coleoptera were identified to at least the family level to assess their potential association with carrion. The following literature was used for identification:
- BAJERLEIN et al. (2012)
- BENISCH (2007)
- BROHMER & SCHAEFER (2018)
- FREUDE et al. (1965), Volumes 4, 8, 13, 15
- LOMPE (2002)
Species identifications were verified using the entomological Fehse Collection held by the Chair of Forest Protection in Tharandt.
Quantification of Diptera Larvae
In addition to the dominant Calliphoridae, other Diptera families such as Sarcophagidae occur on carrion. Undetermined larvae were recorded quantitatively. Freshly hatched L1-stage larvae were not included. It is assumed that most of the larvae belonged to Calliphoridae and Sarcophagidae, although this cannot be confirmed due to the lack of further identification. For large sample sizes (>500–1000 individuals), quantification was performed approximately using volumetric methods (measuring cylinder) in the first trial, and gravimetrically (precision scale) in the second. In both cases, counted subsamples of visually similar larvae served as references. For gravimetric quantification, small batches of approximately 250 larvae were counted for each trap. These were weighed, and the total number of larvae was extrapolated based on the total sample mass and the weight of the reference batch.
Documentation of Additional Arthropods
In addition to Diptera larvae, adult Calliphoridae and Sarcophagidae were collected and their abundance in the traps was recorded. Hymenoptera were also recorded, as they are potential predators of Diptera larvae. Among these, Vespidae and especially Formicidae (ants) played a prominent role (see section 3.3). To evaluate their role in arthropod succession, ants were grouped according to taxonomic and functional characteristics. The subfamily Myrmicinae (e.g., genera of small, less agile ants) was assumed to have little influence on arthropod succession, as their ability to prey on Diptera larvae or fly eggs is likely limited. In contrast, the genus Formica was observed to effectively remove even large L3 Diptera larvae. All other Formicinae (excluding Formica), including Serviformica and Lasius species, were grouped together.
Additionally, Blattodea (cockroaches) were recorded as a potentially necrophagous insect order.
Statistics, Data Analysis, and Visualization
Descriptive analyses and data processing were carried out using Microsoft Excel (Office 365, Microsoft Corporation) and the statistical software RStudio (Version 4.1.2). Data visualization was performed using the RStudio package ggplot. The capture data per carcass were treated as statistically independent. This assumes that the carcasses within each trial period did not interact or lead to species exchange between sites, even though the distances between carcasses were likely surmountable for necrophilous insects. Such interactions are difficult to quantify and were therefore disregarded. For the capture and count data, interpreted as a sample from a large population, a Poisson distribution was assumed, as the probability of any single capture event was relatively low. Rare species occurrences were considered statistical artifacts but were also treated under a Poisson distribution to ensure data comparability (SCHLICHT, pers. comm.).
To examine whether collected taxa showed preferences for specific carcass regions—and thus specific trap positions a Chi-squared goodness-of-fit test was applied. This test is suitable for Poisson-distributed data and is used to assess whether the observed distribution deviates from an expected one. In this study, the null hypothesis stated that the data were evenly (normally) distributed across all trap locations. The analysis focused exclusively on the inner ring of traps (A–D).
If the significance threshold (α) of 5% (α = 0.05) was undercut, the null hypothesis (H₀) was rejected, indicating a non-random distribution, i.e., a preference for or avoidance of specific trap locations by the collected taxa. If sample sizes were small and trap counts did not differ markedly, the null hypothesis could not be rejected. Only taxa or groups with more than ten individuals per carcass were evaluated. It should be noted that the Chi-squared test assumes independence among data points. Since Diptera larvae and Formicidae do not move independently of each other, results for these groups may be biased.
To compare species composition across trial periods, the RENKONEN dominance index (also known as the percent-similarity index) was used. This index accounts for the relative abundances or dominance values of species within a community or habitat. To calculate the index, the smaller dominance value (d₁, d₂, …, dₙ) of each species between the two carcasses is summed. If a species is present only at one site, it is counted with a dominance value of zero (GRUSCHWITZ 1983; SMITH & SMITH 2009).
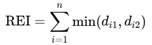
Explanation of the terms:

This index allows for an assessment of the similarity in community composition based on relative species dominance.
Results
General results
Climate Conditions
The climate throughout the year 2021, as recorded by the Grillenburg weather station, was generally consistent with the long-term averages from the reference period 1981–2010. The mean annual air temperature was 7.8 °C, and total annual precipitation amounted to 901.6 mm (Chair of Meteorology 2022) (see Figure 4).
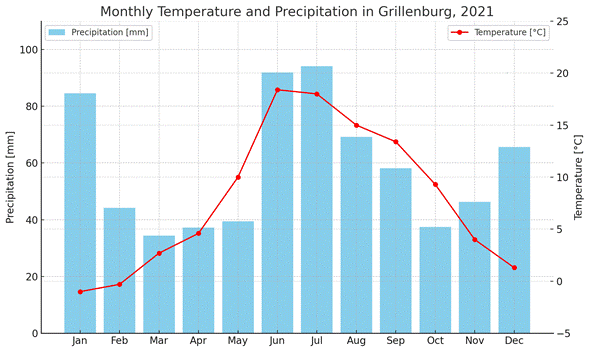
Figure 5: Climate diagram showing monthly values from the Grillenburg weather station, based on data from the Chair of Meteorology (2022).
In April, sub-zero temperatures were recorded on 20 days. Both April (4.6 °C, ΔT = –2.4 °C) and May (10.0 °C, ΔT = –2.3 °C) were cooler than the long-term average. In contrast, June was significantly warmer than average with a mean temperature of 18.4 °C (ΔT = +3.4 °C) (ibid.). At the start of the pitfall trapping in the winter experiment, a sharp temperature increase occurred. While the air temperature at 07:00 on 15 February 2021 was still –18.4 °C, it had risen to 18.9 °C by 15:00 on 23 February 2021 (Chair of Meteorology, 23 January 2021, Grillenburg Weather Station; ΔT = 37.3 °C, measured at 2 m above grass surface). During the second experimental period, August (15.0 °C, ΔT = –1.4 °C) was slightly cooler and September (13.4 °C, ΔT = +1.1 °C) slightly warmer than the long-term average. In August, five meteorological summer days (i.e. days with maximum temperatures exceeding 25.0 °C) were recorded (Chair of Meteorology 2022).
The months of January (+166% deviation) and May (+134% deviation) were significantly wetter than the long-term average. Snowfall continued into April. In contrast, March was noticeably drier, with a deviation of –61% from the long-term average. In August, precipitation was significantly higher than the long-term average, with a deviation of +141%. Almost all of this rainfall occurred during the second half of the month.
Temperature and Humidity Measurements at the Carcass Sites
Differences between the microhabitats (see Tables 17–20) were relatively minor. The average temperatures in the coniferous forest sites were slightly lower than those in the deciduous forest. Larger deviations were observed only in the maximum temperature values. Due to measurement errors, average air humidity could only be partially evaluated.
Decomposition Temperature Measurements Beneath the Carcasses
The temperatures in the winter experiment remained roughly parallel at the deciduous carcass site until mid-April, and at the coniferous carcass site until early May. The highest hourly temperatures measured during the winter experiment were 29.1 °C at the deciduous carcass (22 April 2021) and 35.4 °C at the coniferous carcass (11 May 2021). These data indicate that active decomposition, along with increased Diptera larval activity, began later at the coniferous carcass site.
In the summer experiment, initially very low temperatures were measured beneath the carcasses, which were still frozen at the beginning of the trial. At the deciduous carcass, the temperature beneath the carcass rose more rapidly at first. Nevertheless, the temperature at the coniferous carcass reached its peak one day earlier, with a maximum of 45.5 °C on 14 August 2021, before rapidly returning to baseline levels. A slightly lower maximum temperature of 43.8 °C was recorded at the deciduous carcass on 15 August 2021. The data logger at the deciduous carcass failed on 30 August 2021 and was not replaced.
Vertebrate Scavenger Activity at the Deciduous Carcass
Vertebrate scavenger activity at the deciduous carcass was documented between February 4 and May 29, 2021 (see Table 5). During this period, a pine marten (Martes martes L.) repeatedly accessed the experimental setup and fed on the carcass. In contrast, a raccoon (Procyon lotor L.) was recorded by the camera on only four occasions and did not enter the wire enclosure.
On February 4 and February 23, scavenging was evident without corresponding photographic documentation of the pine marten. However, as it was the only vertebrate scavenger positively identified at the carcass site and was observed frequently, the scavenging events on these two dates are attributed to the pine marten. The absence of photographic evidence is likely due to the camera trap settings at the time; the device was subsequently reconfigured and reinstalled. Until April 2, scavenging by the pine marten was limited to the hind limb musculature, the rumen, and later the dorsal musculature, with the exception of one ear. Between April 2 and April 9, most of the remaining internal organs, including parts of the posterior rib cage, were removed and consumed, leaving only fragments of the rumen and intestines. On April 13, the marten’s scavenging activity exposed the scapula and the hematoma from the bullet wound, which still appeared to be only minimally decomposed (see Figure 30). Additionally, the pine marten marked the carcass on February 23 and March 29 by defecating on the pelvic region. The coniferous carcass was also visited by a raccoon (Procyon lotor L.) (see Table 6). As with the deciduous carcass, the raccoon remained near the carcass only briefly and did not enter the wire enclosure at any time. Given the proximity of the two carcass locations, it is likely that the same individual raccoon visited both sites.
During the summer trial, none of the four camera traps recorded any vertebrate scavengers feeding on the carcasses. However, red deer (Cervus elaphus L.) occasionally triggered the cameras, which rules out a technical malfunction of the equipment.
Temporal Progression of Carcass Decomposition and Associated Arthropods
The decomposition of a carcass begins with the death of the organism and essentially ends with the remaining dry skeletal elements (GOFF 2009). Unlike classical ecological succession patterns, carrion succession does not culminate in a climax community due to resource limitation and spatial constraints (BRAACK 1987, BENBOW et al. 2019). The suitability of the substrate for the organisms involved changes over the course of decomposition (BARTON et al. 2020). Consequently, each stage is characterized by specific groups of arthropods (PAYNE 1965). For the present study, five distinct decomposition stages or succession phases were defined, following conventions common in carrion ecology and forensic analysis. The delineation of these stages is primarily based on physical descriptions found in the literature, although terminology may vary (cf. REED 1958, PAYNE 1965, PAYNE et al. 1968, EARLY & GOFF 1986, TULLIS & GOFF 1987, WANG et al. 2008, GOFF 2009, MATUSZEWSKI et al. 2010a).
The following stage designations are used in this study:
I Fresh Stage
II Bloat Stage
III Active Decay (Decomposition Phase)
IV Dry Decay Phase
V Skeletal Stage
The use of the term stage for stages I and II reflects their relatively well-defined temporal boundaries. In contrast, phases III and IV are referred to as phases to account for the more gradual and subjective nature of their onset and termination. Stage V is again designated as a stage since it marks the cessation of active colonization by necrophilous arthropods, representing a decomposition endpoint—even though the complete depletion of resources may not occur until months or years later.
The transitions between stages are not discrete; rather, they form a continuum (PAYNE 1965, GOFF 2009). Therefore, assigning the exact start and end times of each stage involves a degree of subjectivity.
I Fresh Stage
The fresh stage (Fig. 6) is brief and ends with visible bloating of the abdominal region. Other notable changes include the development of livor mortis on the underside of the body and the onset of rigor mortis. Livor mortis occurs after the cessation of blood circulation, as gravity causes blood to settle in the lower parts of the body. This process typically ends within 9 to 12 hours (GOFF 2009) and is difficult to observe in wild animals due to dense fur. Rigor mortis results from biochemical changes in the muscles: ATP is converted to ADP, and lactic acid lowers the pH in muscle tissue, leading to actin–myosin binding and subsequent stiffening. This stiffening usually persists for one to four days (GOFF 2009). Analyses of odor profiles have shown that freshly deceased carcasses do not emit volatile compounds that differ significantly from those of living organisms (KALINOVÁ et al. 2009, HOERMANN et al. 2013). Typically, blow flies (Calliphoridae) and flesh flies (Sarcophagidae) colonize the carcass at this stage, laying eggs (Calliphoridae) or larvae (Sarcophagidae) in body openings such as the nostrils, ears, eyes, mouth, anus, and any existing wounds (PAYNE 1965, GOFF 2009, BARTON et al. 2020, BYRD & TOMBERLIN 2020b). Moist and sheltered areas like the genital region or inner thighs are also preferred oviposition sites. The reproductive peak of Diptera usually occurs in the subsequent bloat stage, at which point eggs may even be deposited in the gap between the carcass and the ground once other sites are occupied (MATUSZEWSKI et al. 2010a).

Figure 6: fresh stage of roe deer carcass
II Bloat Stage
During the bloat stage (Fig. 7), anaerobic putrefaction—initially driven by gut bacteria—gains prominence. Putrefaction is a bacterial and enzymatic breakdown of soft tissue components (fat, carbohydrates, and proteins), leading to cellular liquefaction, also known as autolysis (DENT et al. 2004, BENNINGER et al. 2008, ANDERSON 2020, BARTON et al. 2020). This typically begins within 48 to 72 hours postmortem (DENT et al. 2004). The metabolic activity of bacteria produces gases, which accumulate in the abdominal cavity, causing the carcass to inflate significantly—sometimes appearing balloon-like (GOFF 2009). Putrefaction continues until oxygen becomes available in phase III. Diptera larval activity begins, producing a mild ammonia odor and attracting additional blow flies and flesh flies. Larvae initially concentrate near the points of entry. Alkaline fluids seep from body openings and into the soil, disrupting the native soil fauna. Organisms typically associated with the active decay phase (stage III) may already begin to colonize the carcass during bloating (GOFF 2009). In spring, bloating tends to persist longer due to delayed Diptera succession; in cold conditions, it may be entirely absent (MATUSZEWSKI et al. 2010a). The bloat stage also sees the highest taxonomic diversity of insects on the carcass (MATUSZEWSKI et al. 2010b).

Figure 7: Bloating stage of the carcass (leaf litter, summer trial)
III Active Decay (Decomposition Phase)
The active decay phase (Fig. 8) begins when abdominal gases escape, permitting oxygen to enter. This may result from internal pressure buildup, larval feeding, or skin rupture (PAYNE 1965, GOFF 2009, BARTON et al. 2020). This stage is characterized by large masses of Diptera larvae, often spilling onto the soil around the carcass (GOFF 2009, MATUSZEWSKI et al. 2010a). Blow fly larvae (Calliphoridae) play a central role (MATUSZEWSKI et al. 2010a). Ongoing microbial activity and larval feeding can generate significant heat, with temperature differences of up to 50 °C relative to the ambient environment (HEWADIKARAM & GOFF 1991, GOFF 2009). The body becomes deflated and emits strong decomposition odors. As oxygen becomes available, anaerobic processes subside. The altered odor profile attracts additional necrophilous arthropods, particularly Diptera larval predators (HOERMANN et al. 2013). The attraction of species like Nicrophorus vespillo and Nicrophorus vespilloides to carrion odor has been well documented (KALINOVÁ et al. 2009, HOERMANN et al. 2013, 2016). Predatory beetles such as Histeridae and Staphylinidae increasingly arrive at the carcass, while necrophagous Coleoptera reproduce on the remains (GOFF 2009). This stage represents the peak of arthropod biomass—particularly Diptera and Coleoptera (COLIJN 2014). Nutrient influx alters soil chemistry, raising pH values (BUMP et al. 2009, HASLAM & TIBBETT 2009). The end of active decay is marked by the near-complete consumption of muscles and internal organs, leaving mostly dry remains (GOFF 2009). In some cases, Necrodes littoralis larvae may dominate over Calliphorids (MATUSZEWSKI et al. 2010a, HÖRMANN 2021).

Fig 8: Onset of the dry decay phase of the carcass
IV Dry Decay Phase
Following active decay , approximately 15% of the carcass’s original mass typically remains under favorable summer conditions; in spring and autumn, this value may be closer to 20% (MATUSZEWSKI et al. 2010a). The dry decay (Fig. 8 and 9) phase, also referred to as “dry remains,” is difficult to define precisely. It usually begins when only small amounts of muscle tissue persist and ends when most necrophilous arthropods have departed, marking the transition to stage V (REED 1958). This phase is independent of rainfall (REED 1958). Diptera are no longer present in significant numbers. Various Coleopteran taxa occur, depending on the moisture content of the remains. Dermestidae—whose adults may already have arrived in the active decay phase—reproduce during this stage, feeding on dry tissue such as skin, cartilage, and residual muscle (GOFF 2009). The phase ends as necrophilous arthropods disperse due to resource depletion and the original soil fauna reestablishes dominance (PAYNE 1965).

Figure 9: Dry decay phase of the carcass (coniferous litter, winter trial)
V Skeletal Stage
In the skeletal stage (Fig. 10), only hair and bones remain. As carrion resources are depleted, necrophagous arthropods disappear. Soil pH gradually returns to baseline levels and may temporarily drop below it (HASLAM & TIBBETT 2009). Meanwhile, the typical soil fauna returns. This stage may persist for months to years, depending on environmental conditions (GOFF 2009). The condition of the carcass also depends on the season of death. In winter, larvae feed less aggressively near the skin surface, allowing more hair to remain. In summer, most carcasses lose their fur almost entirely (REED 1958). Once all muscle tissue is gone, only keratin- and horn-feeding beetles and their predators remain (COLIJN 2014). GU (2014) documented Trogidae, Dermestidae, Cleridae, and even Geotrupidae on bones during this stage. Resistant structures such as teeth and bones persist the longest and decompose primarily through chemical weathering (DENT et al. 2004). In contrast to the temporally defined stages used here, MATUSZEWSKI et al. (2010a) described decomposition as a mosaic of overlapping microhabitats. To better reflect this complexity, they proposed recording decomposition as a set of parallel processes with their own distinct onset and end times. However, for the sake of clarity and simplification, this approach is not adopted in the present study.

Figure 10: Skeletal remains of the carcass (coniferous litter, summer trial)
Winter Trial (January 15 – June 7, 2021)
Deciduous Carcass
Decomposition progressed very slowly at first due to low winter temperatures. The carcass remained in the fresh stage for approximately 20 days and was repeatedly covered by snow. On February 4, a pine marten opened the body cavity, initiating the active decay stage without a preceding bloated phase. Initial larval colonization by Diptera was observed on March 11. Desiccation advanced more rapidly at this site due to higher sun exposure and lower humidity compared to the coniferous site. Calliphoridae and Sarcophagidae adults appeared by late March. A first aggregated larval mass was documented on April 2. Coleopteran activity, dominated by Oiceoptoma thoracicum and Thanatophilus rugosus, was significant. By mid-April, larval mass increased considerably. The peak of larval feeding occurred in late April and gradually declined in early May. The dry decay phase began on May 2. Decomposition continued until only minor tissue and arthropod activity remained by mid-May. The final carcass weight was 4.7 kg.
Coniferous Carcass
This carcass remained in the fresh stage significantly longer due to persistent cold and moisture. The bloated stage began on February 19 (Tab.2), but was weak and fluctuated with sun exposure. Hair slippage in the head and ventral regions occurred in March. First Diptera larvae appeared on March 29. Active decay began in early May, characterized by large larval masses and intense tissue removal, especially from the head and thoracic region. Decomposition advanced rapidly, and by mid-May, most soft tissues were consumed. The dry stage was reached on May 19, and almost all muscle tissue had been removed. Final skeletal condition was documented in early June, with a remaining weight of 4.2 kg.
Summer Trial (August 5 – September 2/10, 2021)
Deciduous Carcass
The carcass was placed frozen and thawed over approximately two days. Decomposition began rapidly due to high ambient temperatures. The bloated stage was observed on August 7 (Tab. 2), marked by early colonization by Calliphoridae. Active decay started on August 11 with extensive larval activity, especially in the head and ventral regions. Dermestidae and Vespidae also appeared. By mid-August, large parts of the head and thorax were skeletonized. The dry decay phase began on August 17, and by the end of the month, most soft tissue was removed, except for some muscle on the hind limbs. Final disarticulation and sclerotization occurred by September 10, marking the skeletal stage. The final weight was 3.5 kg.
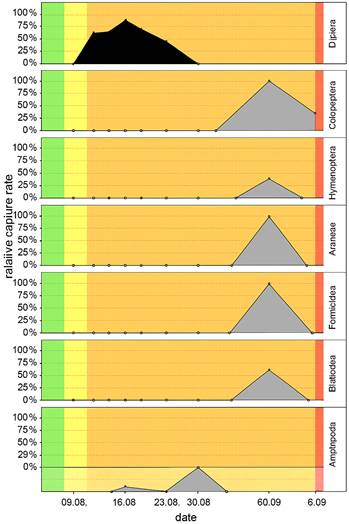

Figure 11: Relative capture rate of Aphodiinae and Scarabaeinae at the leaf litter carcass in the summer experiment.
Coniferous Carcass
The fresh stage lasted until August 7. The bloated stage began on that date and was unusually pronounced, especially in the posterior abdomen. Active decay started on August 11 with concentrated larval activity in the hindquarters. Decomposition progressed rapidly, with high larval density and a strong ammonia odor. Most tissues were consumed by mid-August. The dry decay stage began on August 15. By August 22, few arthropods remained, and on August 27, the carcass reached the skeletal stage. Final weight was 2.9 kg.
Recorded Arthropods – Summary winter
Across both winter trials, a total of 12,696 necrophilous arthropods were collected, including 9,070 Coleoptera. Additionally, 75,286 Diptera larvae were either counted or quantitatively estimated. Only carrion-associated groups were recorded; generalist taxa such as Arachnida, Chilopoda, Collembola, and certain phytophagous and xylophagous Coleoptera (e.g., Elateridae, Curculionidae) were excluded. Typical ectoparasites like Ixodidae and Hippoboscidae were also not considered due to their unrelated ecology.
Table 2: Decomposition stages of the carcasses from the summer experiment
| Carcass | Decomposition stages | Fresh | Bloat stage | Active decay | Dry decay | Skeletal stage |
| Carcass Leaf | 05.08.–07.08.2021 | 07.08.–11.08.2021 | 11.08.–17.08.2021 | 17.08.–10.09.2021 | 10.09.2021 [End of experiment] | |
| Carcass Conifer | 05.08.–07.08.2021 | 07.08.–11.08.2021 | 11.08.–15.08.2021 | 15.08.–27.08.2021 | 27.08.–02.09.2021 [End of experiment] |
Table 3: Decomposition stages of the carcasses from the winter experiment
| Carcass | Fresh | Bloating stage | Active decay | Dry decay | Skeletonization |
| Carcass Leaf litter | 15 Jan – 04 Feb 2021 | / | 04 Feb – 02 May 2021 | 02 May – 07 Jun 2021 | / |
| Carcass Coniferous litter | 15 Jan – 19 Feb 2021 | 19 Feb – 02 May 2021 | 02 May – 19 May 2021 | 19 May – 07 Jun 2021 | / |
Leaf Litter Carcass:
This carcass was opened early by a pine marten, preventing the bloat stage. The early opening led to lower Diptera larval density but prolonged substrate availability for necrophagous beetles.
Common beetles included Oiceoptoma thoracicum, Thanatophilus rugosus, Creophilus maxillosus, and Dermestes murinus. Predatory beetles such as Histeridae and Staphylinidae occurred less frequently.
Diptera larvae: 3,442 counted; adult Calliphoridae and Sarcophagidae were present at moderate density. Formicidae (mainly Formicinae) were consistently active throughout.
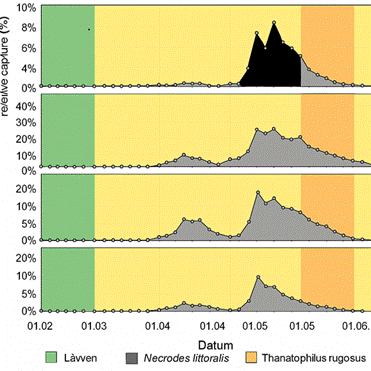

Figure 11a: Abundance as relative capture rate of Silphinae at the deciduous carcass in the winter experiment
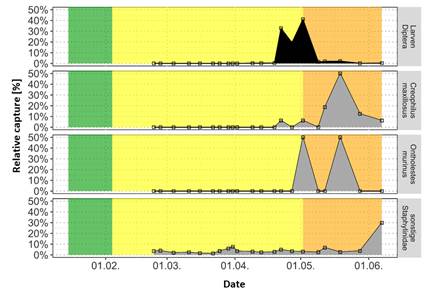
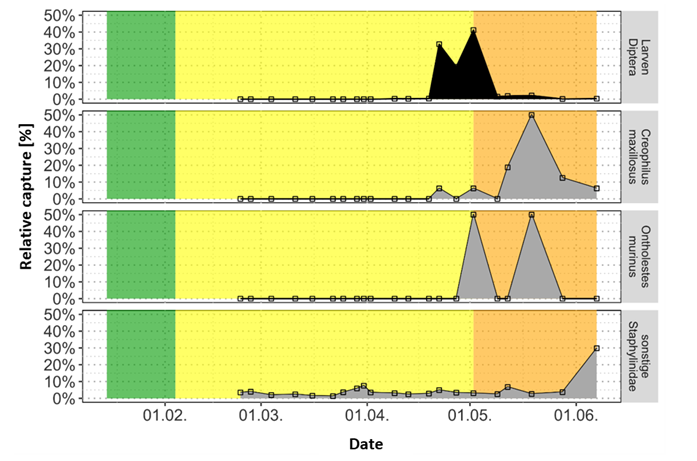
Fig. 11b: Relative catch number of Staphylinidae at the carcass in leaf litter during the winter trial
Conifer Carcass:
Decomposition was delayed due to longer snow cover and lower temperatures.
Diptera larvae were much more abundant (12,850), coinciding with increased numbers of predatory beetles. Necrodes littoralis, Creophilus maxillosus, and Histeridae were much more frequent than at the leaf litter carcass. Oiceoptoma thoracicum and Thanatophilus rugosus showed similar activity patterns but appeared later. Additional dominant groups included Nitidulidae (Omosita depressa), Cholevinae, Ontholestes spp., and Scarabaeidae (A. sticticus).
Comparison (Fig. 12):
Higher Diptera densities at the conifer carcass correlated with increased predator activity. Silphinae were more abundant at the leaf litter carcass; Nicrophorinae were generally rare. Scarabaeidae and Dermestidae dominated at the leaf carcass, while Nitidulidae, Staphylinidae, and Formicinae were more frequent at the conifer site. Peak captures at both carcasses occurred on May 12 and June 7. Overall, capture patterns reflected the course of decomposition and interactions between Diptera and Coleoptera.
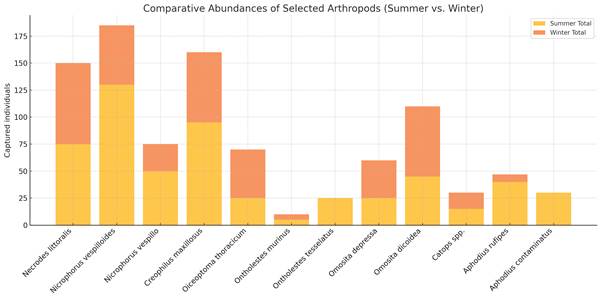
Figure 12: Comparative Abundance of Arthropdes
Recorded Arthropods – Summary summer trial
Despite a similar number of Diptera larvae, beetle abundances differed markedly between the two carcasses (Fig. 13). Predators of Diptera were significantly more abundant at the carcass in leaf litter. This was particularly evident for Necrodes littoralis (6.2×), Creophilus maxillosus (5.3×), and Histeridae (2.4×). Among Silphidae, necrophagous Silphinae species such as Oiceoptoma thoracicum and Thanatophilus rugosus were more frequent at the leaf litter carcass (2.9×), although total numbers were low and presence was short-lived at both sites. Nicrophorinae were more numerous at the leaf litter carcass (258 individuals, 2.5×).
Ontholestes species were present at both carcasses, but more abundant in leaf litter (2.3×), as were other Staphylinidae (1.8×). Cholevinae were more numerous in the needle litter carcass (3.3×), while Nitidulidae were more common in leaf litter (4.1×). Scarabaeidae showed the most pronounced difference: 10 species were recorded in leaf litter versus 5 in needle litter, with Onthophagus spp. occurring exclusively in leaf litter. Scarabaeidae abundance was 11.6× higher in leaf litter. In contrast, more Hydrophilidae were collected from the needle litter carcass (2.4×). Cleridae were present at both carcasses in low numbers, with all three necrophilous Necrobia species found in leaf litter and two in needle litter.
Among Hymenoptera, Vespidae were more frequent at the leaf litter carcass (4.8×). Formica polyctena was virtually absent from the leaf litter carcass but highly abundant (1513 individuals) at the needle litter site, despite the 140 m distance to the nearest nest.
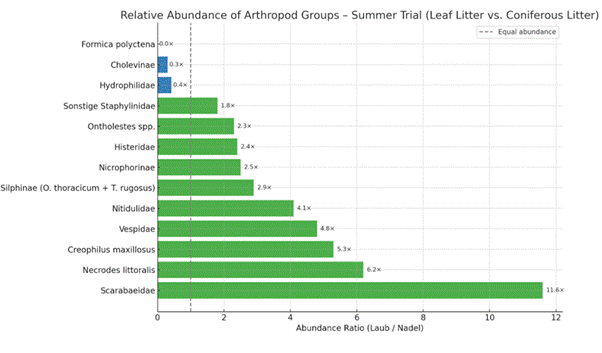
Figure 13: Relative Abundance of Arthropod groups
Seasonal Patterns of Arthropod Abundance in Summer
Silphinae were generally rare, with the exception of N. littoralis at the leaf litter carcass, where high and sustained trap numbers were recorded. At both carcasses, N. littoralis activity followed the abundance of Diptera larvae, though with a delayed decline in the needle litter. O. thoracicum was detected only on a few days and appeared early during bloating at the needle litter carcass. T. sinuatus was also found during the dry decay phase in leaf litter. Nicrophorinae showed strong activity early at the needle litter carcass, peaking on August 9. At both carcasses, N. vespilloides was the first species to appear, but its numbers declined during active decay in needle litter. N. humator was absent during active decay, and N. vespillo showed a brief decline during this stage. C. maxillosus mirrored Diptera larval abundance, with a slight delay at the needle litter site. Ontholestes spp. were rare overall: O. murinus was limited to active decay, O. tesselatus appeared mostly during dry decay, and O. haroldi occurred during dry decay in leaf litter and active decay in needle litter. Other Staphylinidae showed no distinct temporal trend but were continuously present from August 9 onward. Both Omosita species (Nitidulidae) were recorded at both carcasses, with O. dicoidea appearing slightly earlier. In leaf litter, neither species was present at the beginning of dry decay, while in needle litter they were absent on only one day each. Cholevinae were rare in leaf litter. At the needle litter carcass, Catops spp. were absent during active decay and on August 22, but peaked on September 2, suggesting that extended sampling might have provided additional insights. Scarabaeidae were abundant in leaf litter mainly during dry decay. A. sticticus and A. contaminatus peaked on September 2, while A. rufipes peaked on August 22. O. similis appeared earlier, during active decay. In contrast, Scarabaeidae were rare and appeared later in needle litter, with highest numbers on the final sampling day, possibly indicating further increases in the skeletal stage.
Comparison of Abundances Between Seasonal Trials
A comparison between the winter and summer trials reveals notable differences in arthropod abundance and species composition across families. In the winter trial, necrophagous Silphinae—excluding Necrodes littoralis—were dominant within Silphidae, primarily represented by Oiceoptoma thoracicum and Thanatophilus rugosus. The latter was not detected at all during the summer trial. In contrast, the summer trial was characterized by a strong presence of the genus Nicrophorus. Necrodes littoralis occurred at similar frequencies in both trials. Among Nicrophorus species, N. vespilloides was most common in winter, whereas N. vespillo was more abundant overall in summer but less frequent than N. vespilloides at the leaf litter carcass. Within Staphylinidae, fewer Ontholestes species were captured during winter, although the total number of other staphylinids was considerably higher than in summer.Histeridae were more abundant during the summer trial, with Saprinus semistratus dominating in both seasons. In winter, Margarinotus bruneus was also commonly collected. Fewer Cholevinae were recorded in summer, though they were more abundant at the needle litter carcass in both seasons. Nitidulidae were more numerous in winter, with Omosita dicoidea being the most frequent species in both trials. Scarabaeidae were substantially more abundant in winter. Aphodius prodromus dominated winter samples, while the larger A. rufipes was the most frequent species in summer. Hydrophilidae were slightly more common in summer. Cleridae and Dermestidae were rarely recorded in summer; however, three Cleridae species were found only in summer, while three Dermestidae species were present in winter and only one in summer.
For Hymenoptera, numerous Formicinae were captured exclusively in winter. Formica polyctena was abundant only during summer, and only at the needle litter carcass.
Evaluation of Manual Collections
Manual collections revealed that early-occurring, unspecialized groups such as miscellaneous taphy-linidae were frequently recorded at both carcasses during the winter trial. Additionally, earlier occur-rences of Dermestes murinus (needle litter, winter), Oiceoptoma thoracicum (both carcasses, sum-mer), and Creophilus maxillosus (leaf litter, summer) were documented. Manual sampling also enabled later detections of Dermestes frischii (leaf litter, winter), D. murinus (needle litter, summer), and Diptera larvae (needle litter, summer).
Necrobia rufipes (Cleridae) was detected solely through manual sampling at the needle litter carcass in summer. Sampling of Diptera larvae, including from beneath the carcasses, improved documentation of their occurrence.
Necrophilous Arthropods in the Surroundings of the Carcasses
Trap catches at 1 m and 3 m distances from the carcasses were low. For analysis, traps F and G (1 m) and E and H (3 m) were pooled. Since only two pitfall traps were deployed at each distance zone compared to four directly at the carcass (A–D), potential catches are inherently lower. Data (Fig. 13) show that only a few species and a small proportion of individuals occurred beyond the immediate carcass vicinity. In winter, some Silphinae were recorded away from the carcass, including the only capture of Phosphuga atrata at 3 m, suggesting it may not be strongly associated with carrion. Slightly more individuals were recorded in both distance zones around the leaf litter carcass. Miscellaneous Staphylinidae were scarce beyond the inner trap ring and decreased with distance, indicating close carrion association. Tenebrionidae were only captured at 3 m from the leaf litter carcass in summer, suggesting they belong to the general ground beetle fauna rather than being carrion-associated. In the summer trial, only four necrophilous beetle species were found in the surroundings of the needle litter carcass. Despite the overall low beetle abundance at this carcass, it was notable that only Histeridae were found away from the carcass. These occurred later in the decomposition process.
Diptera larvae and Formicidae were highly concentrated at the carcass itself. At 3 m, only 3% of Diptera larvae were captured near the needle litter carcass (summer); all other sites recorded <1%. Slightly higher captures at 1 m and 3 m occurred at this carcass, likely due to mass migration of third instar larvae.
Formicidae also showed carcass-centric distribution, except for Myrmicinae, which were more frequently captured at 1 m and 3 m at three out of four carcasses—suggesting their abundance is not influenced by carrion presence. One exception occurred in the winter trial at the needle litter carcass, where all 32 Formica polyctena individuals were collected in a single trap during one sampling event.
Temporal factors must be considered: abundant species were more frequently recorded at distance, and captures away from the carcass increased after active decay. However, the limited data preclude generalization and may reflect experimental design constraints.
Species Composition Similarity Index
Only beetle taxa identified to species level were considered in assessing differences in species composition. Table 15 presents the number and proportion of identified necrophilous Coleoptera per carcass, serving as the basis for calculating the percent similarity index. The proportion of beetles identified to species level was generally higher in the summer trial compared to the winter trial, and consistently higher at the leaf litter carcass than at the needle litter carcass across both seasons (Fig. 14).
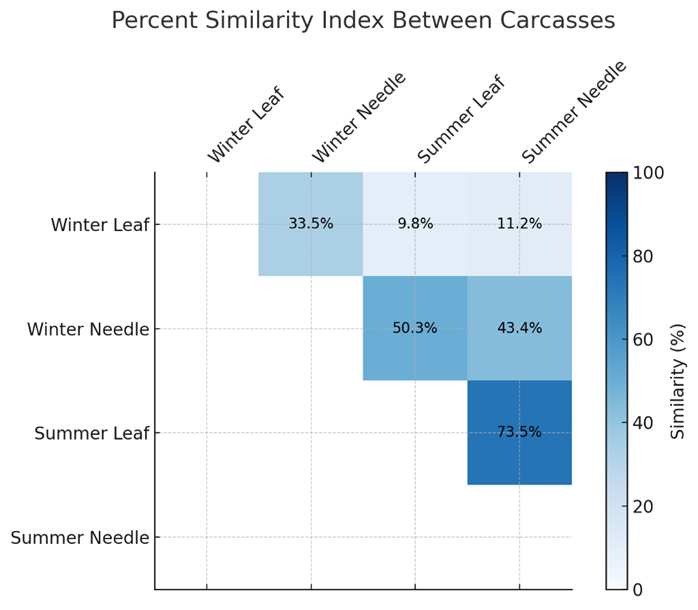
Figure 14: Similary Index of Carcasses
The Renkonen similarity index, calculated from species-level identified individuals and their relative abundances, is presented in Table 16. The analysis indicates that community similarity was generally higher during the summer trial compared to the winter trial. Seasonal comparisons reveal greater variation within the deciduous forest habitat than in the coniferous forest. The index is strongly influenced by the presence of abundant Diptera predators such as Creophilus maxillosus, frequently occurring Histeridae like Saprinus semistratus, and necrophilous Silphidae including Nicrophorus vespilloides and Necrodes littoralis. High similarity values occur when these species are consistently present in large numbers at the compared carcasses.
For frequently collected species (n > 10), a Chi² goodness-of-fit test was applied to assess whether individuals were evenly distributed across the four body regions—head, back, abdomen, and anus—or whether significant deviations indicated preferences or avoidance of specific carcass areas and associated substrates. Corresponding data are provided in Tables 31–38. In cases of significant deviation, the trap with the highest count was highlighted; if the second-highest count exceeded 80% of the maximum, it was also emphasized.
In the winter trial, Dermestidae were most frequently collected in trap C, located at the anal region, at both carcasses. Scarabaeidae, particularly Aphodius spp., were most abundant in trap B (back) during the winter trial at both sites. No other species surpassed this in abundance in any other trap. For the two most frequent species per carcass, the Chi² test confirmed a significant deviation from uniform distribution. At the leaf litter carcass, the second most frequented trap for Scarabaeidae was D (abdomen), whereas no secondary preference was observed at the needle litter carcass. Hydrophilidae at the needle litter carcass were also most frequent in trap B, although the Chi² test did not confirm this pattern as statistically significant. Cleridae were primarily collected in trap A (head) during the winter trial; this distribution approached statistical significance at the leaf litter carcass. Nitidulidae were notably abundant only in traps A and D at the needle litter carcass during winter, while distributions at other carcasses were inconsistent or too sparse for statistical inference. Among necrophagous Silphidae, Oiceoptoma thoracicum and Thanatophilus rugosus showed clear, carcass-specific spatial preferences in winter: most individuals were captured in traps C and D at the leaf litter carcass, and in trap A at the needle litter carcass. Species such as Nicrophorus spp., Creophilus maxillosus, Necrodes littoralis, and Histeridae exhibited spatial distributions similar to that of Diptera larvae. This pattern also applied to Formicidae in the winter trial, with both groups often sharing primary or secondary trap preferences. However, this relationship did not hold at the needle litter carcass in the summer trial.
Discussion and Conclusion
Methodological Considerations
Carcass Selection
Two categories of carcasses were used: hunted roe deer in winter and, in summer, one hunted and one road-killed animal. The winter carcasses were similar in condition, except for a minor weight difference and a shared bullet wound, which did not significantly influence colonization. Contrary to previous reports, the wound site was not preferentially used by Diptera, possibly due to the immediate death and lack of pre-mortem tissue changes. The main factor influencing decomposition was scavenging by a pine marten, which caused premature opening of the torso and altered colonization patterns. In summer, the two carcasses differed more notably. The road-killed deer showed extensive internal damage and retained winter fur, leading to earlier and more intense insect colonization, likely due to injury-related odor cues and potentially advanced decomposition before freezing. Although both carcasses were frozen, a lag in decomposition onset of approximately one day was observed, likely due to incomplete freezing of the road-killed carcass. This emphasizes the need for standardized freezing protocols in comparative decomposition studies. From an ethical standpoint, the use of hunted animals for ecological research aligns with legal and scientific standards, provided the research serves a legitimate scientific purpose.
Experimental Setup
Pitfall traps following Barber’s method proved effective for collecting both diurnal and nocturnal arthropods. However, large numbers of larvae—especially Diptera—were occasionally lost or underrepresented due to trap overflow, disturbance, or obstruction by substrate. Vertebrate scavengers also interfered with some traps despite protective cages, particularly in the winter trial. While vertebrate activity complicates standardization, it reflects natural decomposition scenarios and may increase coleopteran diversity (e.g., dung beetles). Manual sampling (3-minute timed collections) supplemented pitfall data, helping to detect early colonizers and rare species with minimal disturbance. However, variable weather—especially low temperatures—reduced manual collection efficiency in winter. Future studies should consider longer or repeated sampling for improved resolution. Radiation measurements were used to assess habitat comparability. Surprisingly, measured solar input was similar between sites, contradicting initial subjective impressions. This underscores the value of objective microclimatic data in decomposition ecology. Quantifying Diptera larvae was methodologically challenging. A shift from volumetric (winter) to gravimetric (summer) methods improved accuracy. Gravimetric analysis using small sample counts was more efficient and reliable, especially given differences in larval development stages and moisture content. Finally, spacing between carcasses and control sites may have influenced colonization, particularly in summer. Wider spacing (but within practical limits, e.g., <1 km) is recommended to minimize cross-attraction.
Staging of Decomposition
Decomposition stages were defined using a combination of physical changes and arthropod activity. This dual approach accounts for seasonal differences, such as the often indistinct bloating phase in spring. However, stage classification remains somewhat subjective and may differ from other studies due to varying criteria. Species composition can change rapidly, particularly during resource depletion at the end of the dry decay phase. This study found overlaps between late-developing Diptera larvae and early Scarabaeidae activity, suggesting transitional substrate use. Some species (e.g., Diptera) were still detected during the skeletal stage, likely due to delayed pupation and not true ongoing decomposition. Stages were assigned on the day of trap emptying, while trap catches reflect the full preceding interval. This introduces a temporal uncertainty of up to one interval, particularly relevant during rapid summer succession. More frequent sampling would improve accuracy. Future research may benefit from a spatially resolved assessment of decomposition stages at trap locations. For necrophagous beetles, stage assignment by trap position could better reflect the mosaic nature of decomposition. For predatory species, abundance trends could be linked directly to Diptera larval density.
Discussion of Results
Comparison with Other Studies
When comparing the findings of this study with previous carrion ecology research, differences in species abundance and seasonal patterns were observed, though methodological differences limit direct comparability. In open-field studies (e.g., GU 2014), Silphidae were more abundant, particularly Thanatophilus sinuatus, while Nicrophorus species were largely absent. Conversely, the present study documented a greater abundance of Nicrophorus and lower numbers of T. sinuatus, aligning with findings from forest-based studies (e.g., Matuszewski et al. 2010b; Wielink 2004). Notably, Oiceoptoma thoracicum was consistently found in forest habitats, whereas it was absent in open fields. The observed high abundance of Creophilus maxillosus and Necrodes littoralis also matches trends from forest studies, although the timing and frequency of captures varied. The results further support the notion that necrophagous Silphinae dominate in spring, while Nicrophorinae become more prominent in summer. Differences in Histeridae species composition and abundance were also observed, with Margarinotus bruneus being dominant across studies.
The diversity and abundance of Scarabaeidae and Geotrupidae differed significantly between studies, possibly due to habitat, soil properties, and the presence of vertebrate scavengers. Nitidulidae, Dermestidae, and Cleridae patterns were also largely consistent with prior observations, though seasonally variable.
Seasonal Influence
Decomposition progressed more slowly during the winter trial due to low temperatures, resulting in prolonged resource availability and higher diversity, particularly among late colonizers such as Dermestidae and Cleridae. The relative abundance of species and larval development rates of Diptera were delayed, leading to extended successional stages. These findings align with those of Tabor et al. (2004) and Matuszewski et al. (2010a), who documented a seasonal shift in species richness and resource exploitation. The species composition also shifted between seasons: N. humator and N. vespillo appeared early in spring, while N. vespilloides dominated initial colonization in summer. Interestingly, C. maxillosus and N. littoralis exhibited delayed responses to Diptera larval activity, contrary to previous findings (Matuszewski et al. 2011), with higher overall summer activity observed in this study. This suggests season-specific interactions between prey availability and predator response.
Habitat Effects
While habitat-related differences were expected, they were not clearly supported due to limited replication and confounding factors (e.g., vertebrate scavenging, initial carcass condition). Forest type had minor influence on overall colonization, consistent with Matuszewski et al. (2011), who reported larger differences between seasons than between forest habitats. However, variations in soil type (e.g., podzolic brown earth vs. pseudogley) and microclimate may explain differences in Scarabaeidae occurrence. Soil properties likely influence burial behavior and reproduction of dung beetles (e.g., Aphodius spp., Onthophagus), as documented by Hoermann et al. (2020). The occurrence of open-habitat species such as Omosita colon, Dermestes frischii, and Necrobia rufipes may reflect proximity to clearings, although this could not be confirmed. Formicidae abundance reflected microclimatic preferences rather than carrion use. While Formicinae responded to Diptera activity, Myrmicinae appeared largely unrelated to decomposition dynamics. The presence of Formica polyctena in high numbers near a summer carcass was likely due to optimal foraging conditions and nest proximity.
Additional Observations
Capture rates at 1–3 m distance from carcasses were low, likely due to limited vertebrate interference and substrate dispersal. Spatial preferences on the carcass surface showed inconsistent patterns. The expected association of Scarabaeidae with gut content was not supported by trap data. Community similarity analysis (Renkonen index) indicated greater dissimilarity during the winter trial, though differences were more strongly driven by carcass condition and vertebrate activity than by habitat. Future studies should aim for increased replication to differentiate seasonal and habitat-specific effects more robustly. Relative abundance data were influenced by variable sampling intervals. Standardized trap durations or correction factors would improve comparability between intervals.
Limitations
This study is limited by its scale and scope, particularly in terms of species identification. Taxonomic constraints, especially within Staphylinidae, restricted full species-level resolution. Identification challenges are common in carrion ecology, and expert collaboration—as seen in large-scale studies—is often necessary for reliable species lists.The restricted focus on adult Coleoptera and select families was necessary for feasibility but limits generalizability. Comprehensive identification requires broader trapping methods and taxonomic resources, including genitalia preparation for small rove beetles.Temperature data were collected externally due to equipment constraints, limiting conclusions on internal decomposition dynamics. Improved temperature logging would enhance accuracy.Ultimately, this study provides a snapshot of necrophilous beetle communities in two Central European forest habitats and highlights the need for larger, repeated experiments to draw broader ecological or forensic conclusions.
Discussion
Temporal and spatial scales of carcass decomposition and their significance for ecological processes
The decomposition of animal carcasses is a highly dynamic process whose ecological effects extend across different temporal and spatial scales. On a short-term temporal scale (hours to days), carcasses create local hotspots of increased microbial activity, temperature, and nutrient availability (Carter et al. 2007; Finley et al. 2015). These changes lead to the rapid colonization of microorganisms, insects, and vertebrates, resulting in the formation of short-lived but ecologically significant food webs (Barton et al. 2013).
On medium time scales (weeks to months), decomposition has a lasting effect on soil chemistry and microbial communities in the vicinity of the carcass, for example through increased nitrogen and phosphorus concentrations, which manifest themselves in a “cadaver decomposition island” (CDI) (Towne 2000; Parmenter & MacMahon 2009). These islands act as a source of nutrients over long periods of time and can alter local vegetation and soil fauna (Bump et al. 2009).
In terms of spatial scale, it should be noted that, depending on their size and accessibility, carcasses can have effects ranging from a few decimetres (microbial environment, soil fauna) to several kilometers – especially if they are used as a resource by mobile scavengers such as foxes, birds, or bears and are distributed throughout the habitat (Mateo-Tomás et al. 2015). Through these trophic interactions, carcasses can influence material flows and population dynamics beyond their immediate environment.
A comprehensive understanding of the ecological significance of carcasses therefore requires the consideration of multiscale perspectives. The integration of spatial and temporal scales allows both immediate effects on nutrient cycles and longer-term consequences for biodiversity and habitat structure to be captured (Beasley et al. 2012).
Implications of carcass removal measures in protected areas
In many managed landscapes, especially in national parks, forest districts, or nature reserves, the targeted removal of wild animal carcasses is common practice. These measures usually pursue safety, aesthetic, or disease prevention goals (DeVault et al. 2003; Moleón et al. 2014). From an ecological perspective, however, removal can have profound consequences for trophic interactions and material flows. Wildlife carcasses represent a highly concentrated, temporary resource that supports a wide range of organism groups, from microorganisms to invertebrates to large vertebrates (Barton et al. 2013; Bump et al. 2009).
The removal of carcasses can therefore reduce food availability for specialized scavengers (e.g., necrophagous organisms such as Nicrophorus beetles or vultures) and hinder the formation of complex food chains. In the long term, such interventions can impair the biodiversity and functionality of entire ecosystems (Parmenter & MacMahon 2009). In addition, the elimination of natural carrion resources has a negative impact on the population dynamics of obligate scavengers, which is particularly problematic in anthropogenically influenced landscapes (Moleón et al. 2014). Against this background, various authors call for differentiated management that takes ecological processes into account to a greater extent.
Human-wildlife conflicts and disease ecology
However, the presence of animal carcasses in the field can also lead to conflicts with human interests. Particularly near settlements, there is concern that carrion could attract potentially dangerous animals such as bears, wolves, or wild boars, increasing the risk of direct encounters (Selva et al. 2005). In addition, the presence of carcasses is often associated with the spread of zoonotic pathogens or pathogens relevant to animal diseases such as Bacillus anthracis (anthrax) or Mycobacterium bovis (bovine tuberculosis) (Turner et al. 2014; Benavides et al. 2017).
The ecological role of carcasses in pathogen dynamics is complex: while they can potentially act as sources of infection, scavengers also perform an important “sanitary” function in the ecosystem by rapidly removing organic material and thus potentially reducing pathogen persistence (Ogada et al. 2012). Against this background, balanced management that takes both ecological and epidemiological factors into account appears necessary.
Aquatic vs. terrestrial decomposition dynamics: A comparative ecological overview
The decomposition of animal carcasses is a central ecological process that can play fundamentally different functional roles in different habitats. While terrestrial cadaver ecosystems have been increasingly studied and documented in terms of their importance for nutrient cycling, biodiversity, and trophic interactions (Barton et al., 2013; Wilson & Wolkovich, 2011), the comparative perspective on aquatic and terrestrial decomposition systems has been underrepresented to date. An interdisciplinary approach that compares both systems opens up new opportunities to analyze similarities and differences in the control of decomposition processes, nutrient flows, and ecological feedbacks. In terrestrial ecosystems, decomposition is primarily driven by the activity of scavengers (e.g., birds, mammals), detritivores (e.g., insect larvae, worms) and microbial decomposers (e.g., bacteria, fungi) (Carter et al., 2007; Benbow et al., 2019). These organisms interact in complex trophic networks and contribute not only to biomass decomposition but also to the redistribution of energy and nutrients. The release of inorganic compounds such as nitrogen and phosphorus leads to local enrichment of the soil, which promotes so-called “nutrient hotspots,” which in turn attract herbivorous and predatory species (Bump et al., 2009; Parmenter & MacMahon, 2009).In contrast, decomposition in aquatic habitats occurs under physically and chemically very different conditions. Carcasses that enter lakes, rivers, or marine systems are subject to immediate bacterial colonization, often under limited oxygen conditions. While temperature plays a central role, water flow, pH, salinity, and the presence of specialized aquatic detritivores also influence the rate and type of decomposition (Heaton et al., 2020; Haglund & Lindström, 1995). In fast-flowing waters, soluble substances can be transported away quickly, which prevents the concentration of nutrient release in a local area and leads to greater spatial dispersion (Benbow et al., 2015). Another important difference is the availability of scavengers: While terrestrial carcasses are discovered by larger scavengers within a few hours to days, comparable functional groups are absent in many aquatic systems. Here, microorganisms, necrophagous insect larvae (e.g., chironomids) or benthic organisms such as crustaceans play a central role in the decomposition process. This delay or slowing of fragmentation by larger animals can significantly influence the temporal dynamics of decomposition. Studies show that in cold aquatic environments (e.g., deep-sea systems or polar lakes), carcasses can remain intact for months to years before complete decomposition occurs (Haglund & Lindström, 1995). Despite habitat differences, structural similarities can also be identified: In both systems, the presence of carcasses creates a temporary substrate with increased microbial activity, which can lead to an enrichment of local biodiversity (Janzen, 1977; Benbow et al., 2019). The term “necrobiomes” – microbial communities that are specifically specialized to carcass habitats – is relevant for both aquatic and terrestrial systems and can provide clues to functional convergences (Benbow et al., 2015).An integrative comparison of both decomposition worlds also opens up important perspectives for forensic ecology, waste management (e.g., in agriculture and fisheries), and research into nutrient flows in the context of climate change, as changing temperatures and moisture regimes are likely to modulate decomposition differently in both systems.In summary, it can be concluded that the integration of aquatic decomposition systems into cadaver ecology considerations represents an important extension that strengthens the interdisciplinary connectivity of this field of research. A systematic recording of differences in decomposition rates, biological involvement, nutrient retention, and trophic effects provides a basis for comprehensive ecological models and a deeper understanding of ecosystem material flows.
Conflict of interest statement: None declared.
References
- ANDERSON, G. S., (2020): Factors that influence insect succession on carrion. in: Byrd, J. H. & Tomberlin, J. K. [Hrsg.]: Forensic entomology: The utility of arthropods in legal investigations. Third edition. Boca Raton, FL, London, New York, NY: CRC Press Taylor & Francis Group.
- ARCHER, M. S., (2004): Rainfall and temperature effects on the decomposition rate of exposed neonatal remains. in: Science & Justice, Jg. 44, Heft Nr. 1: S. 35-41.
- BAJERLEIN, D.; KONWERSKI, S. und MĄDRA, A., (2012): European beetles of forensic importance: Identification Guide. 21 S. Torun, Poland: EAFE meeting.
- BAJERLEIN, D.; MATUSZEWSKI, S. und KONWERSKI, S., (2011): Insect succession on carrion: Seasonality, habitat preference and residency of histerid beetles (Coleoptera: Histeridae) visiting pig carrion exposed in various forests (Western Poland). in: Polish Journal of Ecology, Jg. 59, Heft Nr. 4: S. 789-797.
- BARBER, H. S., (1931): Traps for cave-inhabiting insects. in: Journal of the Elisha Mitchell Scientific Society, Jg. 46, Heft Nr. 2: S. 259-266.
- BARTON, P. S.; ARCHER, M. S.; QUAGGIOTTO, M.-M. und WALLMAN J. F., (2020): Invertebrate succesion in natural terrestrial enviroments. in: Byrd, J. H. & Tomberlin, J. K. [Hrsg.]: Forensic entomology: The utility of arthropods in legal investigations. Third edition. Boca Raton, FL, London, New York, NY: CRC Press Taylor & Francis Group.
- BARTON, P. S.; CUNNINGHAM, S. A.; LINDENMAYER, D. B. und MANNING, A. D., (2013a): The role of carrion in maintaining biodiversity and ecological processes in terrestrial ecosystems. in: Oecologia, Jg. 171, Heft Nr. 4: S. 761-772.
- BARTON, P. S.; CUNNINGHAM, S. A.; MACDONALD, B. C. T.; MCINTYRE, S.; LINDENMAYER, D. B. und MANNING, A. D., (2013b): Species traits predict assemblage dynamics at ephemeral resource patches created by carrion. in: PloS one, Jg. 8, Heft Nr. 1: S. e53961.
- BARTON, P. S. und EVANS, M. J., (2017): Insect biodiversity meets ecosystem function: differential effects of habitat and insects on carrion decomposition. in: Ecological Entomology, Jg. 42, Heft Nr. 3: S. 364-374.
- Barton, P. S., Cunningham, S. A., Lindenmayer, D. B., & Manning, A. D. (2013). The role of carrion in maintaining biodiversity and ecological processes in terrestrial ecosystems. Oecologia, 171(4), 761–772. https://doi.org/10.1007/s00442-012-2460-3
- BAUCH, T.; ELLIGER, A. und PEGEL, M., (2014): Über 40 Jahre Rehwildmarkierung in Baden Württemberg. in: WFS-Mitteilungen, Jg. 2
- Beasley, J. C., Olson, Z. H., & DeVault, T. L. (2012). Carrion cycling in food webs: comparisons among terrestrial and marine ecosystems. Oikos, 121(7), 1021–1026.
- BENBOW, M. E.; BARTON, P. S.; ULYSHEN, M. D.; BEASLEY, J. C.; DEVAULT, T. L.; STRICKLAND, M. S.; TOMBERLIN, J. K.; JORDAN, H. R. und PECHAL, J. L., (2019): Necrobiome framework for bridging decomposition ecology of autotrophically and heterotrophically derived organic matter. in: Ecological Monographs, Jg. 89, Heft Nr. 1: S. 1-26.
- BENBOW, M. E.; TOMBERLIN, J. K. und TARONE, A. M., (2015): Carrion Ecology, Evolution, and Their Applications. HobokenCRC Press. 596 S.
- Benbow, M. E., Tomberlin, J. K., Tarone, A. M., & Jordan, H. R. (2015). Linking ecosystem services and disservices to necrobiome ecology. Frontiers in Ecology and the Environment, 13(1), 10–17. https://doi.org/10.1890/140179
- Benavides, J. A., Hampson, K., Valderrama, W., Streicker, D. G. (2017). Disease ecology of canine distemper virus in wild carnivores. Animal Conservation, 20(4), 291–302.
- BENISCH, C., (2007): kerbtier.de: Käferfauna Deutschlands. Mannheim. Online im Internet: www.kerbtier.de, Zuletzt geprüft am: 20.1.2022.
- BENNINGER, L. A.; CARTER, D. O. und FORBES, S. L., (2008): The biochemical alteration of soil beneath a decomposing carcass. in: Forensic science international, Jg. 180, Heft Nr. 2-3: S. 70-75.
- BFN, B. F. N., (2021): Einheitlicher Methodenleitfaden “Insektenmonitoring”. 65 S. Bonn
- TierSchG. BMEL, Bundesministerium für Ernährung und Landwirtschaft, (1987): TierSchG – Tierschutzgesetz. 24 S.
- BMU, (2019): Aktionsprogramm Insektenschutz – Gemeinsam wirksam gegen das Insektensterben: Gemeinsam wirksam gegen das Insektensterben, (Bundesministerium für Umwelt, Naturschutz und nukleare Sicherheit, (Hrsg.)). 68 S. Berlin: BMU.
- BONACCI, T.; ZETTO BRANDMAYR, T.; BRANDMAYR, P.; VERCILLO, V. und PORCELLI, F., (2011): Successional patterns of the insect fauna on a pig carcass in southern Italy and the role of Crematogaster scutellaris (Hymenoptera, Formicidae) as a carrion invader. in: Entomological Science, Jg. 14, Heft Nr. 2: S. 125-132.
- BOUREL, B.; MARTIN-BOUYER, L.; HEDOUIN, V.; CAILLIEZ, J. C.; DEROUT, D. und GOSSET, D., (1999): Necrophilous insect succession on rabbit carrion in sand dune habitats in northern France. in: Journal of medical entomology, Jg. 36, Heft Nr. 4: S. 420-425.
- BRAACK, L. E. O., (1987): Community dynamics of carrion-attendant arthropods in tropical african woodland. in: Oecologia, Jg. 72, Heft Nr. 3: S. 402-409.
- BROHMER, P. und SCHAEFER, M., (2018): Brohmer: Fauna von Deutschland: Ein Bestimmungsbuch unserer heimischen Tierwelt. 25., durchgesehene Auflage. WiebelsheimQuelle & Meyer Verlag. 769 S.
- BUMP, J. K.; WEBSTER, C. R.; VUCETICH, J. A.; PETERSON, R. O.; SHIELDS, J. M. und POWERS, M. D., (2009): Ungulate Carcasses Perforate Ecological Filters and Create Biogeochemical Hotspots in Forest Herbaceous Layers Allowing Trees a Competitive Advantage. in: Ecosystems, Jg. 12, Heft Nr. 6: S. 996-1007.
- Bump, J. K., Peterson, R. O., & Vucetich, J. A. (2009). Wolves modulate scavenger behavior in an intact ecosystem. Ecology, 90(4), 891–903.
- BYK, A., (2011): Abundance and composition of Geotrupidae (Coleoptera: Scarabaeoidea) in the developmental cycle of pine stands in Człuchów Forest (NW Poland). in: Baltic Journal of Coleopterology, Jg. 11, Heft Nr. 2: S. 171-186.
- BYRD, J. H. UND TOMBERLIN, J. K. [HRSG.], (2020a): Forensic entomology: The utility of arthropods in legal investigations. Third edition. Boca Raton, FL, London, New York, NYCRC Press Taylor & Francis Group. 585 S.
- BYRD, J. H. und TOMBERLIN, J. K., (2020b): Insect of Forensic Importance. in: Byrd, J. H. & Tomberlin, J. K. [Hrsg.]: Forensic entomology: The utility of arthropods in legal investigations. Third edition. Boca Raton, FL, London, New York, NY: CRC Press Taylor & Francis Group.
- BYRD, J. H. und TOMBERLIN, J. K., (2020c): Laboratory-Rearing of Forensic Insects. in: Byrd, J. H. & Tomberlin, J. K. [Hrsg.]: Forensic entomology: The utility of arthropods in legal investigations. Third edition. Boca Raton, FL, London, New York, NY: CRC Press Taylor & Francis Group.
- CARTER, D. O.; YELLOWLEES, D. und TIBBETT, M., (2007): Cadaver decomposition in terrestrial ecosystems. in: Faculty Publications: Department of Entomology, Jg. 94, Heft Nr. 1: S. 12-24.
- CATTS, E. P. und GOFF, M. L., (1992): Forensic entomology in criminal investigations. in: Annual review of entomology, Jg. 37, Heft Nr. 1: S. 253-272.
- CHIMENO, C.; MORINIÈRE, J.; PODHORNA, J.; HARDULAK, L.; HAUSMANN, A.; RECKEL, F.; GRUNWALD, J. E.; PENNING, R. und HASZPRUNAR, G., (2019): DNA Barcoding in Forensic Entomology – Establishing a DNA Reference Library of Potentially Forensic Relevant Arthropod Species. in: Journal of forensic sciences, Jg. 64, Heft Nr. 2: S. 593-601.
- COE, M., (1978): The decomposition of elephant carcases in the Tsavo (East) National Park, Kenya. in: Journal of Arid Environments, Jg. 1: S. 71-86.
- COLIJN, E. O., (2016): Kevers op kadavers in Nationaal Park De Meinweg (Coleoptera). in: Natuurhistorisch Maandblad, Jg. 105, Heft Nr. 8: S. 154-159.
- COLIJN, O., (2014): Kevers op kadavers in Nederland, de stand van zaken, Jg. 74, Heft Nr. 1-2: S. 60-67.
- DANELL, K.; BERTEAUX, D. und BRÅTHEN, K. A., (2002): Effect of Muskox Carcasses on Nitrogen Concentration in Tundra Vegetation. in: ARCTIC, Jg. 55, Heft Nr. 4
- DEKEIRSSCHIETER, J.; FREDERICK, C.; VERHEGGEN, F. J.; DRUGMAND, D. und HAUBRUGE, E., (2013): Diversity of forensic rove beetles (Coleoptera, Staphylinidae) associated with decaying pig carcass in a forest biotope. in: Journal of forensic sciences, Jg. 58, Heft Nr. 4: S. 1032-1040.
- DEKEIRSSCHIETER, J.; VERHEGGEN, F.; LOGNAY, G. und HAUBRUGE, E., (2011): Large carrion beetles (Coleoptera, Silphidae) in Western Europe: a review, Jg. 15, Heft Nr. 3: S. 435-447.
- DENNO, R. F. und COTHRAN, W. R., (1975): Niche Relationships of a Guild of Necrophagous Flies1,2. in: Annals of the Entomological Society of America, Jg. 68, Heft Nr. 4: S. 741-754.
- DENNO, R. F. und COTHRAN, W. R., (1976): Competitive Interactions and Ecological Strategies of Sarcophagid and Calliphorid Flies Inhabiting Rabbit Carrion. in: Annals of the Entomological Society of America, Jg. 69, Heft Nr. 1: S. 109-113.
- DENT, B. B.; FORBES, S. L. und STUART, B. H., (2004): Review of human decomposition processes in soil. in: Environmental Geology, Jg. 45, Heft Nr. 4: S. 576-585.
- DEVAULT, T. L.; OLSON, Z. H.; BEASLEY, J. C. und RHODES, O. E., (2011): Mesopredators dominate competition for carrion in an agricultural landscape. in: Basic and Applied Ecology, Jg. 12, Heft Nr. 3: S. 268-274.
- DEVAULT, T. L.; RHODES JR., O. E. und SHIVIK, J. A., (2003): Scavenging by vertebrates: behavioral, ecological, and evolutionary perspectives on an important energy transfer pathway in terrestrial ecosystems. in: Oikos, Jg. 102, Heft Nr. 2: S. 225-234.
- EARLY, M. und GOFF, M. L., (1986): Arthropod succession patterns in exposed carrion on the island of O’ahu, Hawaiian Islands, USA. in: Journal of Medical Entomology, Jg. 23, Heft Nr. 5: S. 520-531.
- Finley, S. J., Benbow, M. E., & Javan, G. T. (2015). Microbial communities associated with human decomposition and their potential use as postmortem clocks. International Journal of Legal Medicine, 129(3), 623–632.
- FREUDE, H.; HARDE, K.-W. UND LOHSE, G.-A. [HRSG.], (1965): Die Käfer Mitteleuropas. Spektrum Akademischer Verlag.
- GOFF, L. M., (2009): Early post-mortem changes and stages of decomposition in exposed cadavers. in: Experimental and Applied Acarology, Jg. 49, Heft Nr. 1-2: S. 21-36.
- GOLDBERG, V.; BAUMS, A. UND HÄNTZSCHEL, J. [HRSG.], (2002): Klima, Boden und Landnutzung des Tharandter Waldes: Tharandter Klimaprotokolle. Tharandt
- GOODBROD, J. R. und GOFF, M. L., (1990): Effects of larval population density on rates of development and interactions between two species of Chrysomya (Diptera: Calliphoridae) in laboratory culture. in: Journal of medical entomology, Jg. 27, Heft Nr. 3: S. 338-343.
- GRASSBERGER, M. und FRANK, C., (2004): Initial study of arthropod succession on pig carrion in a central European urban habitat. in: Journal of Medical Entomology, Jg. 41, Heft Nr. 3: S. 511-523.
- GRUSCHWITZ, M., (1983): Populationsökologische Untersuchung zur räumlichen Differenzierung von Carabiden in einem Biotopmosaik (Coleoptera: Carabidae). Bonn
- GU, X., (2014): Animal biodiversity an food web restoration based on large vertebrate carcasses. Brandenburg University of Technology. Cottbus-Senftenberg. 116 S.: Dissertation.
- GU, X.; HAELEWATERS, D.; KRAWCZYNSKI, R.; VANPOUCKE, S.; WAGNER, H.-G. und WIEGLEB, G., (2014): Carcass ecology: more than just beetles. in: Entomologische Berichten, Jg. 74, Heft Nr. 1-2
- GU, X. und KRAWCZYNSKI, R., (2012): Tote Weidetiere – staatlich verhinderte Förderung der Biodiversität. in: Artenschutzreport, Jg. 28: S. 60-64.
- GU, X.; WAGNER, H.-G. und KRAWCZYNSKI, R. Zur Bedeutung toter Großtiere für die Biodiversität: Nationalpark-Jahrbuch Unteres Odertal 2010. S. 21-31.
- GULLAND, F. M. D., (1998): The impact of infectious diseases on wild animal populations: a review. in: Grenfell, B. T. [Hrsg.]: Ecology of infectious diseases in natural populations. Publications of the Newton Institute, 7. Repr. Cambridge: Cambridge Univ. PressS. 20-51.
- Haglund, S. J., & Lindström, J. E. (1995). Decomposition of animal tissue in cold aquatic environments. Journal of Forensic Sciences, 40(2), 276–281.
- Heaton, A. J., Smith, K. E., & Wallman, J. F. (2020). Influence of freshwater habitat on decomposition and insect succession on pig carcasses. Forensic Science International, 307, 110118. https://doi.org/10.1016/j.forsciint.2020.110118
- HASLAM, T. C. F. und TIBBETT, M., (2009): Soils of contrasting pH affect the decomposition of buried mammalian (Ovis aries) skeletal muscle tissue. in: Journal of forensic sciences, Jg. 54, Heft Nr. 4: S. 900-904.
- HESPELER, B., (2016): Rehe in Europa: Biologie und Jagd. WienÖsterreichischer Jagd- und Fischerei-Verlag. 317 S.
- HEWADIKARAM, K. A. und GOFF, M. L., (1991): Effect of carcass size on rate of decomposition and arthropod succession patterns. in: The American journal of forensic medicine and pathology, Jg. 12, Heft Nr. 3: S. 235-240.
- HOERMANN, C. von; JAUCH, D.; KUBOTSCH, C.; REICHEL-JUNG, K.; STEIGER, S. und AYASSE, M., (2018): Effects of abiotic environmental factors and land use on the diversity of carrion-visiting silphid beetles (Coleoptera: Silphidae): A large scale carrion study. in: PloS one, Jg. 13, Heft Nr. 5: S. 1-24.
- HOERMANN, C. von; RUTHER, J. und AYASSE, M., (2016): Volatile Organic Compounds of Decaying Piglet Cadavers Perceived by Nicrophorus vespilloides. in: Journal of Chemical Ecology, Jg. 42, Heft Nr. 8: S. 756-767.
- HOERMANN, C. von; STEIGER, S.; MÜLLER, J. K. und AYASSE, M., (2013): Too fresh is unattractive! The attraction of newly emerged Nicrophorus vespilloides females to odour bouquets of large cadavers at various stages of decomposition. in: PloS one, Jg. 8, Heft Nr. 3: S. 1-11.
- HOERMANN, C. von; WEITHMANN, S.; DEIßLER, M.; AYASSE, M. und STEIGER, S., (2020): Forest habitat parameters influence abundance and diversity of cadaver-visiting dung beetles in Central Europe. in: Royal Society open science, Jg. 7, Heft Nr. 3: S. 1-13.
- HÖRMANN, C., (2021): Beratendes Onlinemeeting zum Versuch und Methodik, [mündlich].
- HOUSTON, D. C., (1986): Scavenging Efficiency of Turkey Vultures in Tropical Forest. in: The Condor, Jg. 88, Heft Nr. 3: S. 318-323.
- HWANG, W., (2006): Konurrenz und Aasnutzung necrophager Käfer in Nord- und Südtaiwan: mit einem Beitrag zur Biologie von Nicrophorus nepalensis (Coleoptera: Siphidae). Albert-Ludwigs-Universität. Freiburg. 110 S.: Dissertation.
- IKEDA, H.; KAGAYA, T.; KUBOTA, K. und ABE, T., (2008): Evolutionary relationships among food habit, loss of flight, and reproductive traits: life-history evolution in the Silphinae (Coleoptera: Silphidae). in: Evolution; international journal of organic evolution, Jg. 62, Heft Nr. 8: S. 2065-2079.
- INSTITUT FÜR WALDSCHUTZ Das Lehr- und Forschungsgehege Grillenburg. TU Dresden, Institut für Waldbau und Waldschutz. Online im Internet: https://tu-dresden.de/bu/umwelt/forst/wb/waldschutz/wildgehege#intro, Zuletzt geprüft am: 26.10.2021.
- JANZEN, D. H., (1977): Why Fruits Rot, Seeds Mold, and Meat Spoils. in: The American Naturalist, Jg. 111, Heft Nr. 980: S. 691-713.
- JOY, J. E.; LIETTE, N. L. und HARRAH, H. L., (2006): Carrion fly (Diptera: Calliphoridae) larval colonization of sunlit and shaded pig carcasses in West Virginia, USA. in: Forensic science international, Jg. 164, Heft Nr. 2-3: S. 183-192.
- KALINOVÁ, B.; PODSKALSKÁ, H.; RŮZICKA, J. und HOSKOVEC, M., (2009): Irresistible bouquet of death: how are burying beetles (Coleoptera: Silphidae: Nicrophorus) attracted by carcasses. in: Die Naturwissenschaften, Jg. 96, Heft Nr. 8: S. 889-899.
- KOČÁREK, P., (2003): Decomposition and Coleoptera succession on exposed carrion of small mammal in Opava, the Czech Republic. in: European Journal of Soil Biology, Jg. 39, Heft Nr. 1: S. 31-45.
- KOREN, T. und TRKOV, D., (2014): The fauna of dung beetles (Scarabaeidae: Aphodiinae, scarabaeinae and geotrupidae) of konavle region, southern croatia. in: Natura Croatica, Jg. 23, Heft Nr. 23 2: S. 389-399.
- KRAWCZYNSKI, R. (2022) Mehr Toleranz für verwesende Leichen. in: Spektrum.de. Online im Internet: https://www.spektrum.de/news/kadaveroekologie-der-wald-braucht-mehr-kadaver/2012221, Zuletzt geprüft am: 27.4.2022.
- KRAWCZYNSKI, R.; GU, X.; RESCHER, S.; LYSAKOWSKI, B. und WIEGLEB, G., (2013): Wirbeltiere an Aas – Erfahrungen aus sechs Jahren Forschung in Brandenburg. in: Jahrbuch Nationalpark Unteres Odertal, Jg. 11: S. 6-15.
- KRAWCZYNSKI, R. und WAGNER, H.-G., (2008): Leben im Tod: Tierkadaver als Schlüsselelemente in Ökosystemen. in: Naturschutz und Landschaftsplanung, Jg. 40, Heft Nr. 9: S. 261-264.
- KRAWCZYNSKI, R. und WAGNER, H.-G., (2010): Ungewöhnliches Nahrungsverhalten der Italienischen Schönschrecke (Calliptamus italicus, L. 1758) und des Heidegrashüpfers (Stenobothrus lineatus, Panzer … in: Articulata, Jg. 25, Heft Nr. 1: S. 23-27.
- KRÜGER, T. und KRÜGER, J.-A., (2007): Einflug von Gänsegeiern Gyps fulvus in Deutschland 2006: Vorkommen, mögliche Ursachen und naturschutzfachliche Konsequenzen. in: Zetischrift für Feldornithologie, Jg. 21, Heft Nr. 3: S. 185-217.
- LFULG und SMEKUL, (2022): Datenportal iDA: interaktive Bodenkarte, Potentielle natürliche Vegetation (pnV). Online im Internet: https://www.umwelt.sachsen.de/umwelt/infosysteme/ida/pages/map/command/index.xhtml;jsessionid=BBE70D7901B8637D609678611B6E044A?mapId=af9d994f-c89a-445a-91dd-cf7816dfd1f8&useMapSrs=true&mapSrs=EPSG%3A25833&mapExtent=394195.7778635117%2C5646121.019399241%2C395799.00145747606%2C5646851.488149241, Zuletzt geprüft am: 4.2.2022, (Freistaat Sachsen, (Hrsg.))
- LOMPE, A., (2002): Käfer Europas: Ein Bestimmungsnetzwerk im Internet. Nienburg
- LUFF, M. L., (1975): Some features influencing the efficiency of pitfall traps. in: Oecologia, Jg. 19, Heft Nr. 4: S. 345-357.
- MALAISE, R., (1937): A new insect-trap. in: Entomologisk tidskrift, Jg. 58, Heft Nr. 1: S. 148-160.
- MARCZAK, D., (2013): Habitat selection by two species of dung beetle, Anoplotrupes stercorosus (Scriba) and Trypocopris vernalis (L.)(Coleoptera: Geotrupidae), changes with stand … in: Forest Research Papers, Jg. 74, Heft Nr. 3: S. 227-232.
- MARTÍN-PIERA, F. und LOBO, J. M., (1996): A comparative discussion of trophic preferences in dung beetle communities. in: Miscel· lània Zoològica, Jg. 19, Heft Nr. 1: S. 13-31.
- Mateo-Tomás, P., Olea, P. P., Moleón, M., Vicente, J., Botella, F., Selva, N., & Sánchez-Zapata, J. A. (2015). From regionalization to integration: Steps and missteps in developing policy for the use of ecosystem services. Ecosystem Services, 16, 1–9.
- MATUSZEWSKI, S.; BAJERLEIN, D.; KONWERSKI, S. und SZPILA, K., (2008): An initial study of insect succession and carrion decomposition in various forest habitats of Central Europe. in: Forensic science international, Jg. 180, Heft Nr. 2-3: S. 61-69.
- MATUSZEWSKI, S.; BAJERLEIN, D.; KONWERSKI, S. und SZPILA, K., (2010a): Insect succession and carrion decomposition in selected forests of Central Europe. Part 1: Pattern and rate of decomposition. in: Forensic science international, Jg. 194, Heft Nr. 1-3: S. 85-93.
- MATUSZEWSKI, S.; BAJERLEIN, D.; KONWERSKI, S. und SZPILA, K., (2010b): Insect succession and carrion decomposition in selected forests of Central Europe. Part 2: Composition and residency patterns of carrion fauna: Composition and residency patterns of carrion fauna. in: Forensic science international, Jg. 195, Heft Nr. 1-3: S. 42-51.
- MATUSZEWSKI, S.; BAJERLEIN, D.; KONWERSKI, S. und SZPILA, K., (2011): Insect succession and carrion decomposition in selected forests of Central Europe.: Part 3: Succession of carrion fauna. in: Forensic science international, Jg. 207, Heft Nr. 1-3: S. 150-163.
- MATUSZEWSKI, S.; SZAFAŁOWICZ, M. und JARMUSZ, M., (2013): Insects colonising carcasses in open and forest habitats of Central Europe: search for indicators of corpse relocation. in: Forensic science international, Jg. 231, Heft Nr. 1-3: S. 234-239.
- MEGYESI, M. S.; NAWROCKI, S. P. und HASKELL, N. H., (2005): Using Accumulated Degree-Days to Estimate the Postmortem Interval from Decomposed Human Remains. in: Journal of Forensic Sciences, Jg. 50, Heft Nr. 3: S. 1-9.
- MELIS, C.; SELVA, N.; TEURLINGS, I.; SKARPE, C.; LINNELL, J. D. C. und ANDERSEN, R., (2007): Soil and vegetation nutrient response to bison carcasses in Białowieża Primeval Forest, Poland. in: Ecological Research, Jg. 22, Heft Nr. 5: S. 807-813.
- MELIS, C.; TEURLINGS, I.; LINNELL, J. D.; ANDERSEN, R. und BORDONI, A., (2004): Influence of a deer carcass on Coleopteran diversity in a Scandinavian boreal forest: a preliminary study. in: European Journal of Wildlife Research, Jg. 50, Heft Nr. 3: S. 146-149.
- Moleón, M., Sánchez-Zapata, J. A., Selva, N., Donázar, J. A., & Owen-Smith, N. (2014). Inter-specific interactions linking predation and scavenging in terrestrial vertebrate assemblages. Biological Reviews, 89(4), 1042–1054.
- NICHOLS, E.; SPECTOR, S.; LOUZADA, J.; LARSEN, T.; AMEZQUITA, S. und FAVILA, M. E., (2008): Ecological functions and ecosystem services provided by Scarabaeinae dung beetles. in: Biological Conservation, Jg. 141, Heft Nr. 6: S. 1461-1474.
- ODDEN, J.; LINNELL, J. D. C. und ANDERSEN, R., (2006): Diet of Eurasian lynx, Lynx lynx, in the boreal forest of southeastern Norway: the relative importance of livestock and hares at low roe deer density. in: European Journal of Wildlife Research, Jg. 52, Heft Nr. 4: S. 237-244.
- Ogada, D. L., Keesing, F., & Virani, M. Z. (2012). Dropping dead: causes and consequences of vulture population declines worldwide. Annals of the New York Academy of Sciences, 1249(1), 57–71.
- ORO, D.; GENOVART, M.; TAVECCHIA, G.; FOWLER, M. S. und MARTÍNEZ-ABRAÍN, A., (2013): Ecological and evolutionary implications of food subsidies from humans. in: Ecology Letters, Jg. 16, Heft Nr. 12: S. 1501-1514.
- PARMENTER, R. R. und MACMAHON, J. A., (2009): Carrion decomposition and nutrient cycling in a semiarid shrub–steppe ecosystem. in: Ecological Monographs, Jg. 79, Heft Nr. 4: S. 637-661.
- PAYNE, J. A., (1965): A Summer Carrion Study of the Baby Pig Sus Scrofa Linnaeus. in: Ecology, Jg. 46, Heft Nr. 5: S. 592-602.
- PAYNE, J. A.; KING, E. W. und BEINHART, G., (1968): Arthropod succession and decomposition of buried pigs. in: Nature, Jg. 219, Heft Nr. 5159: S. 1180-1181.
- PETRUSKA, F., (1975): The effect of predominating winds on the flight of some species of beetles from the group of Silphidae into pitfall traps (Col. Silphidae). in: Acta Universitatis Palackianae Olomucensis, Jg. 51, Heft Nr. 55: S. 175.
- PEWE, T. L.; RIVARD, N. R. und LLANO, G. A., (1959): Mummified seal carcasses in the McMurdo Sound Region, Antarctica. in: Science (New York, N.Y.), Jg. 130, Heft Nr. 3377: S. 716.
- PHILIPS, T. K., (2011): The evolutionary history and diversification of dung beetles. in: Simmons, L. W. & Ridsdill-Smith, J. [Hrsg.]: Ecology and evolution of dung beetles. 1. publ. Oxford: Wiley-BlackwellS. 21-45.
- PRESSESTELLE DES DEUTSCHEN JAGDVERBANDES, (21. Februar 2022): Quellenangabe zur Streckenstatistik des Deutschen Jagdverbandes, [E-Mail].
- PROFESSUR FÜR METEOROLOGIE, (23. Januar 2021): Klimadaten 2021 Station Grillenburg, [E-Mail].
- PROFESSUR FÜR METEOROLOGIE, (2022): Jahresbereicht Klimastation Grillenburg. Tharandt
- RAESFELD, F.; NEUHAUS, A. H. und SCHAICH, K., (1985): Das Rehwild: Naturgeschichte, Hege und Jagd. 14. Aufl. HamburgParey.
- RAY, R.-R.; SEIBOLD, H. und HEURICH, M., (2014): Invertebrates outcompete vertebrate facultative scavengers in simulated lynx kills in the Bavarian Forest National Park, Germany. in: Animal Biodiversity and Conservation, Jg. 37, Heft Nr. 1: S. 77-88.
- REED, H. B., (1958): A Study of Dog Carcass Communities in Tennessee, with Special Reference to the Insects. in: The American Midland Naturalist, Jg. 59, Heft Nr. 1: S. 213-245.
- RITZ, M., (2022): Deutschland | Landkarten kostenlos – Cliparts kostenlos | Seite 4: Landkarte & Stadplanindex. Landkarten und Stadtplan Index. Online im Internet: https://www.landkartenindex.de/kostenlos/?cat=3&paged=4, Zuletzt geprüft am: 9.5.2022.
- ROBERTS, L. G. und DABBS, G. R., (2015): A Taphonomic Study Exploring the Differences in Decomposition Rate and Manner between Frozen and Never Frozen Domestic Pigs (Sus scrofa). in: Journal of forensic sciences, Jg. 60, Heft Nr. 3: S. 588-594.
- SÄCHSISCHES LANDESAMT FÜR UMWELT, LANDWIRTSCHAFT UND GEOLOGIE, (2010): Bodenlehrpfad Tharandter Wald: Exkursionsführer. Dresden
- SÁNCHEZ-BAYO, F. und WYCKHUYS, K. A., (2019): Worldwide decline of the entomofauna: A review of its drivers. in: Biological Conservation, Jg. 232: S. 8-27.
- Selva, N., Jędrzejewska, B., Jędrzejewski, W., & Wajrak, A. (2005). Factors affecting carcass use by a guild of scavengers in European temperate woodland. Canadian Journal of Zoology, 83(12), 1590–1601.
- SCHLICHT, R., (mündlich): Konsultation,
- SCHOENLY, K. und REID, W., (1983): Community structure of carrion arthropods in the Chihuahuan Desert. in: Journal of Arid Environments, Jg. 6, Heft Nr. 3: S. 253-263.
- SCHOENLY, K. und REID, W., (1987): Dynamics of heterotrophic succession in carrion arthropod assemblages:: discrete seres or a continuum of change? in: Oecologia, Jg. 73, Heft Nr. 2: S. 192-202.
- SCHWALBE, E.; MAAS, H.-G.; KENTER, M. und WAGNER, S., (2009): Hemispheric Image Modeling and Analysis Techniques for Solar Radiation Determination in Forest Ecosystems. in: Photogrammetric Engineering & Remote Sensing, Jg. 75, Heft Nr. 4: S. 375-384.
- SCOTT, M. P., (1998): The ecology and behavior of burying beetles. in: Annual review of entomology, Jg. 43, Heft Nr. 1: S. 595-618.
- SEBASTIÁN-GONZÁLEZ, E.; BARBOSA, J. M.; PÉREZ-GARCÍA, J. M.; MORALES-REYES, Z.; BOTELLA, F.; OLEA, P. P.; MATEO-TOMÁS, P.; MOLEÓN, M.; HIRALDO, F.; ARRONDO, E.; DONÁZAR, J. A.; CORTÉS-AVIZANDA, A.; SELVA, N.; LAMBERTUCCI, S. A.; BHATTACHARJEE, A.; BREWER, A.; ANADÓN, J. D.; ABERNETHY, E.; RHODES, O. E.; TURNER, K.; BEASLEY, J. C.; DEVAULT, T. L.; ORDIZ, A.; WIKENROS, C.; ZIMMERMANN, B.; WABAKKEN, P.; WILMERS, C. C.; SMITH, J. A.; KENDALL, C. J.; OGADA, D.; BUECHLEY, E. R.; FREHNER, E.; ALLEN, M. L.; WITTMER, H. U.; BUTLER, J. R. A.; DU TOIT, J. T.; READ, J.; WILSON, D.; JERINA, K.; KROFEL, M.; KOSTECKE, R.; INGER, R.; SAMSON, A.; NAVES-ALEGRE, L. und SÁNCHEZ-ZAPATA, J. A., (2019): Scavenging in the Anthropocene: Human impact drives vertebrate scavenger species richness at a global scale. in: Global change biology, Jg. 25, Heft Nr. 9: S. 1-13.
- SEIFERT, B., (2007): Die Ameisen Mittel- und Nordeuropas. Tauerlutra Verlag. 368 S.
- SELVA, N.; JĘDRZEJEWSKA, B.; JĘDRZEJEWSKI, W. und WAJRAK, A., (2005): Factors affecting carcass use by a guild of scavengers in European temperate woodland. in: Canadian Journal of Zoology, Jg. 83, Heft Nr. 12: S. 1590-1601.
- SHARANOWSKI, B.; WALKER, E. G. und ANDERSON, G. S., (2008): Insect succession and decomposition patterns on shaded and sunlit carrion in Saskatchewan in three different seasons. in: Forensic science international, Jg. 179, Heft Nr. 2-3: S. 219-240.
- SIMMONS, L. W. UND RIDSDILL-SMITH, J. [HRSG.], (2011): Ecology and evolution of dung beetles. 1. publ. OxfordWiley-Blackwell. 347 S.
- SIMMONS, L. W. und RIDSILL-SMITH, T. J., (2011): Reproductive competition and its impact on the evolution and ecology of dung beetles. in: Simmons, L. W. & Ridsdill-Smith, J. [Hrsg.]: Ecology and evolution of dung beetles. 1. publ. Oxford: Wiley-BlackwellS. 1-20.
- SMEKUL [HRSG.], (2022): Waldzustandsbericht 2021. in: Sonderheft. GraupaStaatsbetrieb Sachsenforst.
- SMITH, T. M. und SMITH, R. L., (2009): Ökologie. Pearson Studium Bio-BiologieDt. Ausg., 6., aktualisierte Aufl. [der amerikan. Ausg.]. MünchenPearson Education. 982 S.
- SÖNNICHSEN, L.; BOROWIK, T.; PODGÓRSKI, T.; PLIS, K.; BERGER, A. und JĘDRZEJEWSKA, B., (2017): Survival rates and causes of mortality of roe deer Capreolus capreolus in a rural landscape, eastern Poland. in: Mammal Research, Jg. 62, Heft Nr. 2: S. 141-147.
- STEVENS, J. R.; PICARD C. J. und WELLS J. D., (2020): Molecular genetic methods for forensic entomology. in: Byrd, J. H. & Tomberlin, J. K. [Hrsg.]: Forensic entomology: The utility of arthropods in legal investigations. Third edition. Boca Raton, FL, London, New York, NY: CRC Press Taylor & Francis Group.
- STIEGLER, J.; HOERMANN, C.; MÜLLER, J.; BENBOW, M. E. und HEURICH, M., (2020): Carcass provisioning for scavenger conservation in a temperate forest ecosystem. in: Ecosphere, Jg. 11, Heft Nr. 4: S. 1-13.
- STUBBE, C., (2008): Rehwild: Biologie, Ökologie, Hege und Jagd. 5., neu bearb. Aufl., Neuausg. StuttgartKosmos. 398 S.
- TABOR, K. L.; BREWSTER, C. C. und FELL, R. D., (2004): Analysis of the successional patterns of insects on carrion in southwest Virginia. in: Journal of medical entomology, Jg. 41, Heft Nr. 4: S. 785-795.
- TEURLINGS, I. J. M.; MELIS, C.; SKARPE, C. und LINNELL, J. D. C., (2020a): Lack of Cascading Effects of Eurasian Lynx Predation on Roe Deer to Soil and Plant Nutrients. in: Diversity, Jg. 12, Heft Nr. 9: S. 352.
- TEURLINGS, I. J. M.; ODDEN, J.; LINNELL, J. D. C. und MELIS, C., (2020b): Caching Behavior of Large Prey by Eurasian Lynx: Quantifying the Anti-Scavenging Benefits. in: Diversity, Jg. 12, Heft Nr. 9: S. 1-9.
- Towne, E. G. (2000). Prairie vegetation and soil nutrient responses to ungulate carcasses. Oecologia, 122(2), 232–239.
- TULLIS, K. und GOFF, M. L., (1987): Arthropod succession in exposed carrion in a tropical rainforest on O’ahu Island, Hawai’i. in: Journal of Medical Entomology, Jg. 24, Heft Nr. 3: S. 332-339.
- TUMER, A. R.; KARACAOGLU, E.; NAMLI, A.; KETEN, A.; FARASAT, S.; AKCAN, R.; SERT, O. und ODABAŞI, A. B., (2013): Effects of different types of soil on decomposition: an experimental study. in: Legal medicine (Tokyo, Japan), Jg. 15, Heft Nr. 3: S. 149-156.
- Turner, W. C., Kausrud, K. L., & Stenseth, N. C. (2014). Lethal exposure: factors influencing carcass visitation by vertebrate scavengers in an African savanna. Ecosphere, 5(10), 1–14.
- VÖLLGER, E., (1985): Fangmethoden für Bremsen. in: Entomologische Nachrichten und Berichte, Jg. 29, Heft Nr. 1: S. 91-95.
- WANG, J.; LI, Z.; CHEN, Y.; CHEN, Q. und YIN, X., (2008): The succession and development of insects on pig carcasses and their significances in estimating PMI in south China. in: Forensic science international, Jg. 179, Heft Nr. 1: S. 11-18.
- WEITHMANN, S.; HOERMANN, C. von; SCHMITT, T.; STEIGER, S. und AYASSE, M., (2020): The Attraction of the Dung Beetle Anoplotrupes stercorosus (Coleoptera: Geotrupidae) to Volatiles from Vertebrate Cadavers. in: Insects, Jg. 11, Heft Nr. 8: S. 1-16.
- WIELINK, P., (2004): Kadavers in de kaaistoep:: de natuurlijke successie van kevers en andere insecten in een vos en een ree. in: Entomologische Berichten, Jg. 64, Heft Nr. 2: S. 34-50.
- WILMERS, C. C. und GETZ, W. M., (2004): Simulating the effects of wolf-elk population dynamics on resource flow to scavengers. in: Ecological Modelling, Jg. 177, Heft Nr. 1-2: S. 193-208.
- WILSON, E. E. und WOLKOVICH, E. M., (2011): Scavenging: how carnivores and carrion structure communities. in: Trends in Ecology & Evolution, Jg. 26, Heft Nr. 3: S. 1-7.
- WOLF, J. M. und GIBBS, J., (2004): Silphids in urban forests: Diversity and function. in: Urban Ecosystems, Jg. 7, Heft Nr. 4: S. 371-384.
- WULF, C., (2020): Den Menschen neu denken im Anthropozän. in: Paragrana, Jg. 29, Heft Nr. 1: S. 13-35.
- YOUNG, T. P., (1994): Natural Die-Offs of Large Mammals: Implications for Conservation. in: Conservation Biology, Jg. 8, Heft Nr. 2: S. 410-418.
- ZEILER, H., (2009): Rehe im Wald. WienÖsterriechischer Jagd- und Fischerei Verlag.